We wish to thank Jamie Poston and Christopher Shults for their assistance in conducting this experiment.
Address correspondence to Robert F. Simons, Department of Psychology, University of Delaware, Newark, DE 19716.
![]()
Abstract
Consistent and specific relationships exist between two emotion dimensions (valence and arousal) and self-report, physiological and overt-behavioral responses. These relationships, most frequently studied by varying stimulus content, can be modulated by the form (e.g. screen size, color) in which the stimulus is presented. The present experiment focused on another important presentation attribute -- stimulus motion, and explored its impact on the perceived quality and physiological sequelae of the induced emotion.
Moving and still versions of emotion-eliciting stimuli were delivered to subjects while facial-muscle, heart-rate, skin-conductance activity, and emotion self-reports were monitored. During both moving and still images, facial-muscle and heart-rate change was associated with stimulus valence while skin conductance was associated with arousal. The impact of motion was dramatic. Self-report and physiological data strongly suggested that motion increased arousal, had little impact on valence and captured and sustained the subject's attention to the image.
During the course of the past decade or so, there has been a resurgence of interest in the specific relationships between emotion and its physiological sequelae. Whether emotion is viewed in terms of discrete entities (e.g. Ekman, 1989; Izard, 1997, Tomkins, 1962; 1963) or as a multidimensional space (e.g. Russell, 1997; Russell & Mehrabian, 1977; Lang, Bradley & Cuthbert, 1997), an overarching goal for many psychophysiological investigators is the profiling of each emotion or emotion dimension by determining the pattern of physiological changes with which these emotion variables are associated. From their discrete-emotion perspective and using their directed facial action task, Ekman, Levenson, Ekman & Friesen (1983; Levenson, Ekman & Friesen, 1990), have, for example, demonstrated that facial expressions indicative of specific emotions such as fear, anger, sadness and joy can be associated with specific patterns of autonomic activation. Similarly, Lang (e.g. Greenwald, Cook & Lang, 1989) and others (Fitzgibbons & Simons, 1992; Fiorito & Simons, 1994) have shown that specific relationships exist between facial muscle activity, heart rate and emotion valence ratings and between emotion arousal ratings and the frequency and amplitude of skin conductance responses.
In all of these studies, it was the specific content of the stimulus that was manipulated, either in imagery or with color slides or film clips. Though these studies have produced very interesting data, they have ignored a second aspect of the stimulus materials that might also contribute to the emotion-response topography. This 'non-content' aspect is the form in which the stimulus is delivered. These structural attributes of the stimulus-delivery system are being widely studied by communication researchers and these studies have shown that variables such as screen size (Detenber & Reeves, 1996), viewing distance (Lombard, 1995) and color (Sherman & Dominick, 1988) can have a significant impact on how subjects experience and evaluate stimulus content. It is likely that these same formal message properties can influence the physiological aspect of the emotion response as well.
We (Detenber, Simons & Bennett, 1998) recently reported our first study in which a formal presentation attribute served as the independent variable. In this study, we chose to examine whether the emotion message (image) presented to the subjects was moving or still. We chose to study motion because it is central to most media presentations and is thought to be particularly potent in emotion elicitation (Arnheim, 1983, Gianetti, 1976). Moving images are also more memorable (Kipper, 1986) and are associated with greater cortical activation (Reeves, Thorson, Rothschild, McDonald, Hirsch & Goldstein, 1985). Reeves et al. (1985) suggest that this greater cortical arousal associated with motion is indicative of motion's ability to automatically capture attention.
In our study, 18 undergraduates were shown two versions of each of 27 images. One version was still, the second was moving. Selection of image content was guided by the dimensional view of emotion such that one-third of the images were positively valent, one-third were neutral and one-third were negative. Images were presented for six seconds during which heart rate and skin conductance were measured. Subjects rated each image for valence, arousal and interest shortly after image termination.
The results of this study were intriguing. Image motion had a significant impact on self-reports of arousal and on the magnitude of the skin conductance response -- particularly those associated with images rated high on arousal. Image motion had a much smaller (though still significant) impact on valence ratings and had no impact on the valence-sensitive acceleratory component of the heart-rate response. There was suggestive evidence, however, that other aspects of the heart-rate response were modulated by image motion, but this effect was independent of heart rate's covariation with valence. Motion appeared to produce a deceleration that was sustained throughout the six-second presentation interval; the heart-rate response to the still images began its return to baseline well before stimulus offset.
While these results confirmed the potency of image motion, both on self-report and on physiological measures, they also raised a number of questions that need to be further clarified. First, there is the question of specificity. Are the effects of image motion specific to the arousal dimension of emotion? A specific motion/arousal relationship is consistent with both the memory (moving images are better remembered) and cortical activation (moving images prompt greater desynchrony) data described above. That is, high-arousal images are more accurately recalled than low arousal images while image valence is independent of memory (Bradley, Greenwald, Petry & Lang, 1992) and cortical desynchrony is generally thought to index activation or arousal. In our study, subjects reported moving images to be more arousing than still images, and although they did report that moving images were generally more positive also, the effect on valence was smaller than the effect of motion on arousal reports. In addition, the two physiological measures were consistent with dimensional specificity -- that is, heart-rate acceleration (valence) was not affected by motion but skin conductance (arousal) was. The heart rate data are somewhat ambiguous, however. Heart rate was influenced by motion, though late in the interval and valence effects on heart rate have proven, in general, to be fairly weak relative to some other valence-sensitive measures (e.g. facial EMG).
The present study was designed to examine in more detail the relationship between image motion and emotion response topography. It is clear from our previous study that motion acts to heighten the arousal value of an image. It is less clear how, or whether,the valence of the image. Though we reported a significant enhancement of valence self-report (SAM), there was no effect of motion on valence self report in a previous study by Detenber and Reeves (1996), and our heart-rate data were ambiguous. The purpose of the present study, therefore, was to confirm the significant effect of image motion of arousal related measures (SAM, SCR) while bringing more power to bear on the relationship between motion and valence. This was accomplished by incorporating into the procedure, the assesssment of facial muscles known to be sensitive to stimulus valence (i.e. corrugator and zygomatic) and by enlarging the sample size in order to provide a more sensitive test of heart-rate change -- particularly the late-interval deceleration that seemed to characterize the response to images containing motion. It was our hypothesis that effects of motion would be observed on arousal- but not valence-related measures and that motion's effect on heart-rate activity would be distinct from the relationship between heart rate and image valence.
![]()
Subjects
Thirty-five undergraduate students at the University of Delaware participated in this study and received partial credit toward the research participation component of their introductory psychology course or extra credit in their mass communication course. Of the original 35 subjects, 1 subject discontinued due to illness, and data from two additional subjects were eliminated from heart-rate analyses, three from the skin conductance analysis and 3 from the EMG analyses due to technical problems during data collection. The final sample of 34 consisted of 17 malesand 17 females with a mean age of 19.24 years (sd=2.35).
Stimuli
The stimuli consisted of 27 images sampled from films and television programs. The images chosen for use in the present study were a subset of those used previously by Detenber (1995) and identical to those used by Detenber, Simons and Bennett (in preparation). The selection of images was based upon categories appearing in the International Affective Picture System (IAPS; Lang, Ohman, & Vaitl, 1988) and these particular 27 images were associated with a wide range of ratings on the emotion dimensions of interest in the present study (i.e. valence and arousal).
All stimuli were presented for 6 s and were either moving or still versions of the same image. The still version of each image was simply one of the frames taken from the full motion clip that was highly representative. All images, along with an additional image instructing subjects to perform the ratings task, were stored on a video laser disc that was connected to a Macintosh computer. Stimuli were presented to subjects in one of four orders embedded in a Hypercard control program used by the Macintosh to control the sequence and the timing of stimulus presentation.
Response measurement
Self-report
. Subjects provided ratings of valence, arousal and dominance for each of the 54 images by completing a nine-point paper and pencil version of Lang's Self-Assessment Manikin (SAM; Lang, 1980). Using the SAM, valence is rated by marking on or between five graphics depicting the manikin with facial expressions ranging from a broad smile to a severe frown. Arousal is rated similarly using five graphics depicting the manikin at different levels of visceral agitation and dominance is rated using manikins which differ in size or prominence in the graphics panel they occupy.Physiological recording
. Heart-rate was obtained by attaching a Grass Photoelectric Transducer Model PPS to the subject's right ear lobe. The photocell output was fed into a Grass Model 7P1 Low Level DC Preamplifier and Model 7D Driver Amplifier (Bandpass = 1.6 - 3.0 Hz) and then into a Grass Model 7P4 Cardiotachometer where the interpulse intervals were converted into heart rate in beats per minute (BPM).Skin conductance responses were recorded using a Coulbourn Model S21-22 constant voltage (.5V) skin conductance coupler. Prior to recording, the palm of the nonpreferred hand was cleansed with distilled water. Beckman Standard (0.5 cm2) Ag/AgCl electrodes were then placed on the thenar and hypothenar eminence of the palm with Johnson and Johnson KY Jelly used as electrolyte.
Electromyographic (EMG) recordings from the face were obtained by placing Med-Associates miniature Ag/AgCl electrodes over the subject's left zygomatic and corrugator muscles. The raw EMG (Bandpass = .15 - 500 Hz) was full-wave rectified and integrated (TC=50 ms) using a Grass Model 7P3 Wideband Amplifier/Integrator.
Procedure
Subjects were provided with a brief description of the stimuli, the ratings task and the recording techniques and then signed an informed consent form. EMG and skin conductance electrodes were then affixed on their respective recording sites and the subject was led to an adjacent room equipped with a comfortable arm chair positioned approximately 1.4 m in front of the viewing device (SONY 23" Color Monitor). The photocell was attached to the ear and the quality of the physiological recordings was inspected. Subjects then received the complete set of instructions and two 'neutral' practice trials were delivered. The experiment began if the instructions were understood, if the ratings task was completed properly during the practice trials and if the physiological recordings were free of obvious noise and artifact.
The experiment proper consisted of 54 trials under the control of two laboratory computers -- a 486 PC that initiated each trial and collected the physiological data and the Macintosh that controlled the laser disc player. Each trial began with a signal, through a simple serial connection, from the PC to the Macintosh. The signal caused the Macintosh to deliver one of the fifty four images for 6 s. For 1 s following the completion of the clip, the viewing screen was dark, and then the instruction to rate the image was presented on screen for 4 s. Subjects were instructed to rate the image on the three dimensions (valence, arousal, dominance) quickly, and to return their eyes to the viewing screen prior to the appearance of the next image. The period for ratings varied randomly from 17 to 27 s. Physiological data collection began 2 s prior to the delivery of each image and continued for 10 s. At the half-way point in the experiment, the experimenter reentered the viewing room to provide a short break for the subject and to ensure that the subject was on the appropriate page in the ratings booklet. At the conclusion of the experiment, subjects were verbally debriefed and given a brief written explanation of the experiment along with some relevant citations.
Data Reduction
The skin conductance, cardiotachometer, and two facial EMG channels were sampled at 50 cps. The skin conductance data were displayed graphically, trial by trial, and quantified by visually identifying response onset and the largest peak that occurred with an onset latency of 0.5 - 4 s following stimulus onset. SCR magnitude was defined as the difference, in uSiemens, between the identified peak and onset points.
The cardiotachometer data were edited for artifact by visually inspecting each trial. Bad beats were generally replaced by the trial average. If, however, the baseline heart rate was bad or if consecutive bad beats were detected during image presentation, the entire trial was deleted and omitted from the appropriate condition average. Heart rate waveforms were obtained from the edited cardiotachometer record by averaging successive 25 data points, and deviating each half-second average during the 8 s post-onset epoch from the half-second average immediately preceding stimulus onset. Fourteen (7 s) half-second averages, along with the onset point, constituted the heart-rate data that were then submitted to statistical analysis.
Corrugator and zygomatic muscle activity was also examined on each trial. Trials with unstable baseline periods were deleted. Trials with obvious artifacts during the viewing period were edited (mean substitution) wherever possible, or deleted. Corrugator and zygomatic data were then expressed as the difference between the mean value during the 6 s viewing period and the prestimulus mean. A derived 'pattern' score was also computed for each trial by subtracting the corrugator change from the zygomatic change (Fitzgibbons & Simons, 1992; Greenwald, Cook & Lang, 1989).
Data Analysis
The initial phase of data analysis involved the generation of mean valence and arousal ratings for each of the images collapsed across the moving/still dimension. Valence means were then ranked from most positive to least positive and then the 27-image set was divided into 9 positive, 9 neutral and 9 negative images. Likewise, arousal means were ranked from the lowest to highest and the images were divided into 9 low-, medium- and high-arousal categories, again collapsed across the moving/still dimension.
Each of the dependent measures was analyzed twice using a repeated-measures Analysis of Variance (ANOVA) with Image Category (Valence or Arousal) and Motion as the two within-subject variables. Orthogonal trends were used to represent the Category variable. In this analysis, the linear trend (1,0,-1) is equivalent to the specific contrast of positive v negative valence or low v. high arousal, whereas the quadratic trend (1, -2, 1) is equivalent to the contrast of the middle category with the two extremes. The heart rate analysis also examined orthogonal trends across the half-second data points to assess the effects of both image Category and Motion across time.
![]()
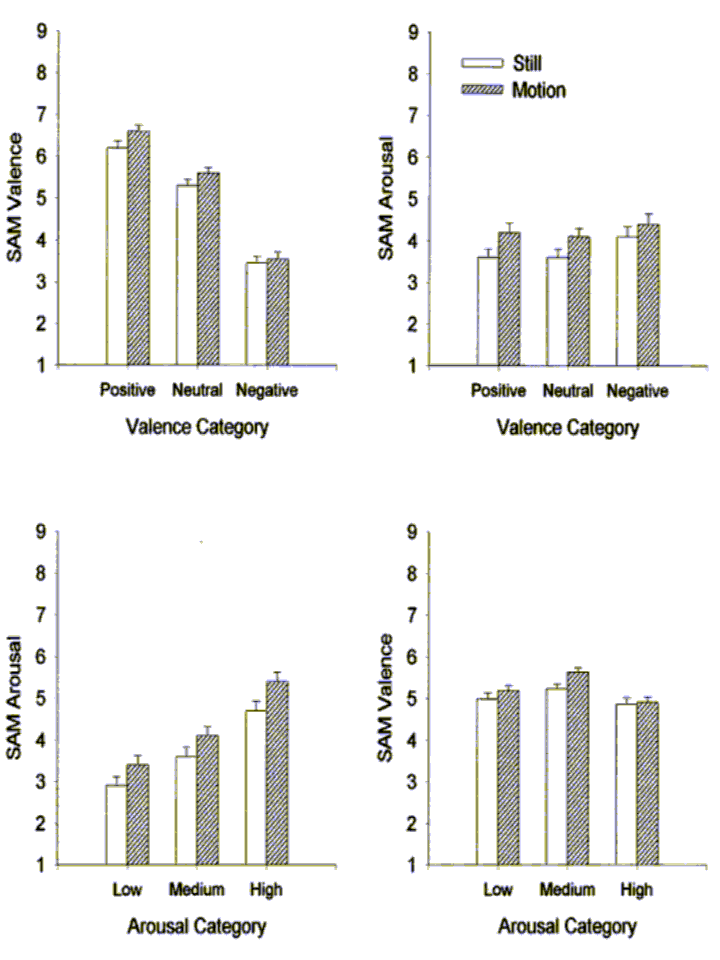
SAM Ratings
Valence and arousal ratings as a function of both valence and arousal categories are presented in Figure 1. The two left-hand panels illustrate the impact of image motion on SAM ratings of valence (top) and arousal (bottom). Moving images were rated more positively than still images overall (F(1,33)=18.89, p<.001) and motion interacted with linear valence (Flin(1,33)=11.57, p<.01); the difference between moving and still images grew as valence became more positive. Likewise, the effect of motion on arousal ratings was highly significant with moving images experienced as more arousing than still images (F(1,33)=54.54, p<.001).
The two right-hand panels of Figure 1 illustrate the relationship between valence and arousal ratings. The relationship between valence category and SAM arousal ratings was quadratic with positive and negative slides rated as more arousing than the neutral slides (Fquad(1,33)=9.97, p<.01). Likewise, the relationship between arousal category and SAM valence ratings was also quadratic. In this case, slides of medium arousal were rated more pleasant than high and low arousal slides (Fquad(1,33)=33.5, p<.001). The linear correlation between ratings of valence and arousal in the present study was nil (r=.009).
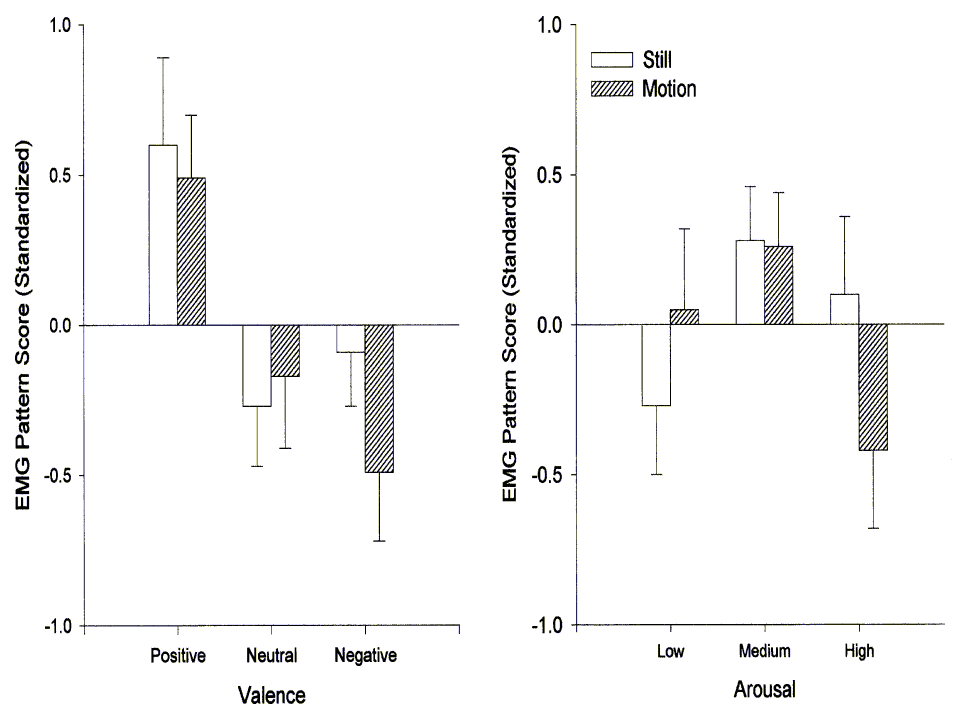
Facial EMG
Figure 2 illustrates the relationship between the facial EMG pattern score and both valence and arousal categories. As expected, the pattern score varied linearly with image valence (Flin(1,30)=11.36, p<.01). This linear trend was evident in both of the constituent facial muscles (zygomatic Flin(1,30)=4.24, p<.05; corrugator Flin(1,30)=9.14, p<.01) with positive images prompting greater zygomatic and negative images greater corugator activity. Somewhat surprisingly, the EMG pattern score also varied with arousal category. This relationship was significantly quadratic (Fquad(1,30)=7.52, p<.05) with both low- and high-arousal slides evincing a corrugator-type pattern. This pattern was exclusively due, in fact, to activity in the corrugator (zygomatic Fquad(1,30)=1.52, ns; corrugator Fquad(1,30)=6.01, p<.05). Motion did not exert a main effect on facial EMG and it did not interact with image valence. As Figure 2 suggests, however, motion did interact significantly with the arousal category (Flin(1,30)=7.48, p<.05). This interaction can be accounted for by a substantial corrugator response to highly arousing moving images that was absent in response to high-arousal stills (zygomatic Flin(1,30)<1; corrugator Flin(1,30)=5.41, p<.05).
Heart Rate
The heart-rate response to the image stimuli was deceleratory, beginning shortly after stimulus presentation and remaining below baseline for the duration of the recording interval. The half-second by half-second data are presented in Figure 3 as a function of image valence (left-hand panel), image arousal (center panel) and image motion (right-hand panel). The trend analysis of variance confirmed the reliability of the deceleration with significant linear (Flin(1,31)=41.36, p<.001), quadratic (Fquad(1,31)=36.88, p<.001) and quartic (Fquart(1,31)=11.80, p<.01) trends across the presentation period. The linear and quadratic trends accounted for 77% and 17% of the variance across the half-second data points respectively.
Heart-rate change was significantly related to both emotion properties of the stimuli (i.e. valence and arousal). The relationship between stimulus valence and heart rate was linear (Flin(1,31)=8.33, p<.05; Fquad(1,31)<1) with the greatest deceleration associated with negative images and the least with the positive images. The development of this between-image differentiation across time is reflected in the significant valence X linear half-second interaction (Flin(1,31)=15.43, p<.001). The arousal properties of the image were similarly related to heart-rate change, though in this case, the relationship was both linear (Flin(1,31)=5.63, p<.05) and quadratic (Fquad(1,31)=6.88, p<.05) reflecting the greater deceleration associated with the most highly arousing images and the lack of differentiation between stimuli rated low and medium in arousal. The development of this association over time was again supported by an interaction of stimulus category (arousal) with half-second time points (Flin(1,31)=7.98, p<.01; Fquad(1,31)=7.76, p<.01). Full differentiation of the stimuli by both emotion dimensions was evident in the heart-rate response by mid-interval and remained stable until the image was removed.
Motion also had a significant impact on the heart-rate response (F(1,31)=10.78, p<.01) with moving images prompting more heart-rate slowing than the still images. As Figure 3 illustrates, the impact of motion also developed across time (Motion X linear Half-second Flin(1,31)=29.36, p<.001). In this case, however, the differentiation increased throughout the presentation period and was greatest at the end of the period, rather than at mid-interval. Motion exerted its effects on heart rate independently; there were no interactions between motion and either of the two emotion dimensions.
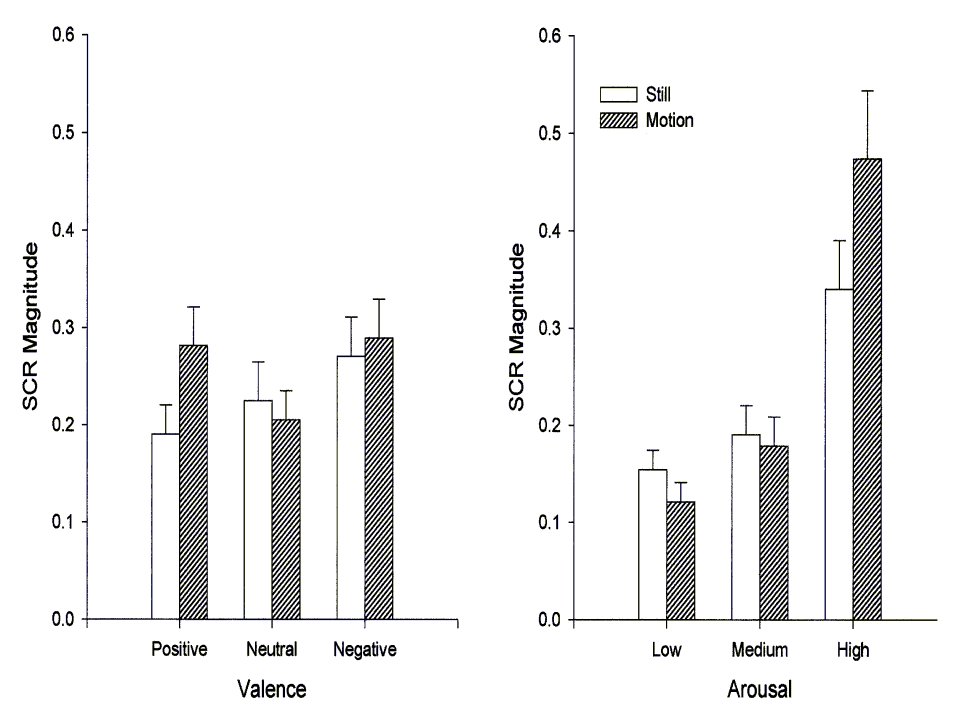
Skin Conductance
As expected, skin conductance response magnitude was a function of the arousal properties of the image stimuli. This relationship was primarily linear, accounting for 87% of the arousal-category variance, though the quadratic was also significant
(Flin(1,30)=25.39, p<.001; Fquad(1,30)=12.31, p<.01). As the right-hand panel of Figure 4 illustrates, high-arousal images evoked particularly large skin conductance responses. The figure also illustrates the highly significant association between image motion and SCR magnitude. Statistically, motion interacted with arousal by accentuating the linear trend (Motion X linear Arousal Flin(1,30)=12.31, p<.01). The left-hand panel of Figure 4 depicts the relationship between SCR magnitude and image valence. The relationship was significantly quadratic (Fquad(1,30)=7.01, p<.05), most likely reflecting the fact that both positive and negative images tend to be more arousing than the neutral images. Motion did not interact with valence category.
![]()
The main goal of the present study was to confirm the Detenber et al. (1998) finding that the impact of motion in an emotion-evoking image is primarily on variables associated with emotion’s arousal dimension. This we did successfully. Moving images were once again associated with increases in self-reported arousal and with larger skin conductance responses, particularly when the images were reported to be highly arousing. Motion also prompted a substantial and sustained bradycardia suggesting that moving images captured more attentional resources.
As in Detenber et al. (1998), there was also a significant effect of stimulus motion on valence self-report. This effect, however, was much smaller than the arousal effect and did not extend to either the valence-sensitive heart-rate acceleration measure or to facial EMG – perhaps the prototypic nonverbal index of emotional valence. Although both facial EMG and heart-rate acceleration were unequivocally associated with image valence, as expected, these effects were independent of the motion/still manipulation.
Together, the effects reported by Detenber et al. (1998) and those observed in the present study suggest that image motion, an attribute of stimulus presentation independent of content, can have a powerful impact on the experience and physiology of emotion and that this impact can be rather specific. That is, image motion moderates the arousal value of the image to a greater extent than it moderates the subjects’ like or dislike of image content. A similar specificity has been found in studies of screen size; Detenber and Reeves (1996) reported arousal, not valence self-report increases when images are presented on large viewing screens and, more recently, Reeves, Lang, Kim and Tatar (1997) have shown that the screen size effect is evident in skin conductance as well as arousal self-reports.
A second major goal of the present experiment was to provide a more complete explication of the difference in the heart-rate response to the moving and still images noted by Detenber et al (1998). As discussed above, heart rate is reliably associated with stimulus valence in picture viewing and in other emotion-eliciting tasks. This relationship was obvious in the present data as well. Recall how the heart-rate waveforms clearly distinguished the positive, neutral and negative images independent of the motion factor (Figure 3; left). Likewise, image motion exerted a strong effect on the heart-rate response and this effect was independent of stimulus valence. Movement in the image was associated with marked deceleration, and this deceleratory difference between moving and still images grew most pronounced toward the end of the viewing period (Figure 3; right).
Based on these data, it appears that as the heart-rate response unfolds, several processes are reflected, not unlike those described by Bohlin & Kjellberg (1979). First, all stimuli elicit a short-latency deceleration indicative of orienting to image onset (Graham, 1992; Graham & Clifton, 1966). Next, the affective properties of the stimulus dominate and their impact is most evident around the mid-interval acceleratory component. Finally, motion becomes the dominant factor. Heart rate returns to baseline if the image is still, but remains substantially slowed if the image contains motion. This continued bradycardia most likely reflects sustained attention (e.g., Berg & Richards, 1997; Richards, 1997). Thus, it appears that the ‘message’ (i.e., stimulus content) and the ‘medium’ both affect the heart-rate response and, in this case at least, these effects are independent.
This interpretation of the heart-rate data is consistent with other studies of formal media characteristics and their physiological effects. In general, the onset of new or novel visual information in a mediated presentation has been associated with heart-rate deceleration. For example, Thorson and Lang (1992) found that the introduction of a videographic (i.e., text on the screen) in a televised lecture caused a significant slowing in the viewers’ heart rate for several seconds. Similarly, it has been shown that edits in television program segments that involve a change of scene elicit cardiac orienting responses (Lang, Geiger, Strickwerda, & Sumner, 1993). Lang (1990) presented evidence that both the onset and novel structural features (i.e., edits, camera motion, object motion onscreen, etc.) of commercials embedded in a half-hour television program can cause heart-rate decelerations. What distinguishes our data from the data of Lang and her colleagues is the sustained slowing of the heart. That is, there appears to be an effect of motion beyond simple orienting. We believe that motion, by definition, continues to present new information to viewers, and thereby holds their attention once it has been captured by content.
The present study, then, provides clear evidence that how a stimulus is delivered affects emotion-response topography above and beyond the effects of which stimulus is delivered. Furthermore, the effects of presentation attributes such as motion and screen size can be fairly specific. Both of the latter effects increase arousal and sustain the subjects attention (see Reeves et al., 1997, for a more complete discussion of screen size effects) while having little or no impact on image valence. These data have implications for research in the psychology of emotions as well as for the field of communication. Most media effects research examines only the content of messages and its impact. Based on our results, media-effects researchers would do well to consider the potential impact of the form of the messages and the possibility of form and content interactions. Media producers concern themselves with message effectiveness. Often, the criterion for effectiveness is the ability to capture and hold the audience’s attention. Our studies indicate that motion can play a critical role in eliciting and sustaining attention. Motion can also increase emotional arousal or excitement, and this is often another goal of media producers.
Emotion research, likewise, has focused primarily on stimulus content and has paid little attention to the formal properties of stimulus presentation. Given the goal of inducing emotional responses in the laboratory that mimic, as much as possible, those that occur 'in vivo', it is important to identify manipulations that may affect the potency of the emotion-inducing stimuli. The present study, and those of Reeves and his colleagues (e.g. Detenber & Reeves, 1996; Reeves et al., 1997) have identified two variables, image motion and screen size, that augment the potency of the stimuli. In both cases, the impact of the variable has been to make the stimuli more arousing. Perhaps other presentation attributes (e.g. color, sound?) might be identified that increase the potency of a stimulus by affecting more specifically its valence -- i.e., making positive images more positive and negative images more negative. Independent manipulations of valence and arousal through formal presentation properties would have interesting and important implications for basic research in emotion and for research in disordered emotion as well. We have noted, for example, that subjects with physical anhedonia have emotion-processing deficits that are primarily valence related (Fitzgibbons & Simons, 1992; Fiorito & Simons, 1994) and have found more recently that subjects identified as alexithymic have, what appear to be, arousal-related deficits (Roedema & Simons, in press). Such an analysis might prove useful with other subject groups as well, and the ability to identify, and then target manipulations toward specific emotion dimensions could have theoretical and practical implications that are both interesting and important.
![]()
Arnheim, R. (1958/1983). Film as art. London: Faber & Faber.
Berg, W.K. & Richards, J.E. (1997). Attention across time in infant development. In P.J. Lang, R.F. Simons, & M.T. Balaban (Eds.), Attention and orienting: Sensory and motivational processes (pp. 347-368). Hillsdale, NJ: Lawrence Erlbaum Associates.
Bohlin, G. & Kjellberg, A. (1979). Orienting activity in two-stimulus paradigms as reflected in heart rate. In H.D. Kimmel, E.H. van Olst, & J.H. Orlebeke (Eds.), The orienting response in humans (pp. 167-197). Hillsdale, NJ: Lawrence Erlbaum Associates.
Bradley, M., Greenwald, M.K., Petry, M.C. & Lang, P.J. (1992). Remembering pictures: Pleasure and arousal in memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 18, 379-390.
Detenber, B. H., & Reeves, B. (1996). A bio-informational theory of emotion: Motion and image size effects on viewers. Journal of Communication, 46(3),66-84.
Detenber, B.H., Simons, R.F. & Bennett, G.G. (1998). Roll 'Em: The effects of picture motion on emotional responses. Journal of Broadcasting and Electronic Media, 42, 112-126.
Ekman, P. (1989). The argument and evidence about universals in facial expressions of emotion. In H. Wagner & A. Manstead (Eds.) Handbook of Social Psychophysiology (pp.143-164). Chichester, England: Wiley & Sons.
Ekman, P., Levenson, R.W. & Friesen, W.V. (1983). Autonomic nervous system activity distinguishes among emotions. Science, 221, 1208-1210.
Fitzgibbons, L. & Simons, R.F. (1992). Affective response to color-sllde stimuli in subjects with physical anhedonia: A three-systems analysis. Psychophysiology, 29, 613-620.
Fiorito, E.R. & Simons, R.F. (1994). Emotional imagery and physical anhedonia. Psychophysiology. 31, 513-521.
Gianetti, L.D. (1976). Understanding movies (2nd ed.). Englewood Cliffs, NJ: Prentice Hall.
Graham, F. K. (1992). Attention: The heartbeat, the blink, and the brain. In B. A. Campbell, H. Hayne, & R. Richardson (Eds.), Attention and information processing in infants and adults (pp. 3-29). Hillsdale, N.J., Lawrence Erlbaum.
Graham, F.K. & Clifton, R.K. (1966). Heart rate change as a component of the orienting response. Psychological Bulletin, 65, 305-320.
Greenwald, M.K., Cook, E.W. & Lang, P.J. (1989). Affective judgment and psychophysiological response: Dimensional covariation in the evaluation of pictorial stimuli. Journal of Psychophysiology, 3, 51-64.
Izard, C.E. (1997). Emotions and facial expressions: A perspective from differential emotions theory. In J.A. Russell & J.M. Fernandez-Dols (Eds.) The psychology of facial expression (pp 57-77). New York: Cambridge University Press.
Kipper, P. (1986). Television camera movement as a source of perceptual information. Journal of Broadcasting & Electronic Media, 30, 295-307.
Lang, P.J., Bradley, M. & Cuthbert, B.N. (1997). Motivated attention: Affect, activation and action. In P.J. Lang, R.F. Simons & M.T. Balaban (Eds.) Attention and orienting: Sensory and motivational processes (Pp. 97-136). Mahway, NJ: Lawrence Erlbaum Associates.
Levenson, R.W., Ekman, P. & Friesen, W.V. (1990). Voluntary facial action generates emotion-specific autonomic nervous system activity. Psychophysiology, 27, 363-384. Lombard, M. (1995). Direct responses to people on the screen: Television and personal space. Communication Research, 22, 288-324.
Reeves, B., Lang, A., Kim, E.Y. & Tatar, D. (May, 1997). The effects of screen size and message content on attention and arousal. Paper presented to the Information Systems Division of the International Communication Association, Montreal, Canada.
Reeves, B.E. Thorson, E., Rothschild, M., McDonald, D., Hirsch, J. & Goldstein, R. (1985). Attention to television: Intrastimulus effects of movement and scene changes on alpha variation over time. International Journal of Neuroscience, 25, 241-255.
Richards, J.E. (1997). Effects of attention on infants' preference for briefly exposed visual stimuli in the paired-comparison recognition-memory paradigm. Developmental Psychology, 33, 22-31.
Russell, James A. (1997). How shall an emotion be called? In R. Plutchik & H.R. Conte (Eds.) Circumplex models of personality and emotions (p. 205-220). Washington, DC: American Psychological Association.
Russell, James A. & Mehrabian, A. (1977). Evidence for a three-factor theory of emotions. Journal of Research in Personality, 11, 273-294. Sherman, B. L., & Dominick, J. R. (1988). Perceptions of colorization. Journalism Quarterly, 65, 976-980.
Tomkins, S. S. (1962). Affect, imagery, consciousness: Vol I. The positive affects. New York: Springer.
Tomkins, S. S. (1963). Affect, imagery, consciousness: Vol II. The negative affects. New York: Springer.
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}