Preparation of this manuscript was supported, in part, by a Biomedical Research Grant from the University of Delaware to the second author. Portions of this research were submitted by the first author to the University of Delaware in partial fulfillment of the requirements for the Doctor of Philosophy degree. The Ph.D. was awarded posthumously to Dr. Fiorito who died on November 26, 1992 following a long illness. This manuscript is submitted in honor of her extraordinary courage and of the fond memory in which she is held by all her friends and colleagues.
Address correspondence to Robert F. Simons, Department of Psychology, University of Delaware, Newark, DE 19716.
![]()
A previous study by Fitzgibbons and Simons (1992) confirmed that subjects identified as anhedonic consistently report a reduced experience of pleasure when confronted with hedonic stimuli (i.e. color slides). Evidence that this emotion-processing deficit extended from the verbal report to the physiological and behavioral domains was mixed, but suggested that deficient imagery ability might be associated with the reduced capacity to experience pleasure. The present experiment was designed to test this hypothesis explicitly.
Anhedonic and normal control subjects were presented scripts which they were to imagine as vividly as possible. During both script presentation and imagery, heart rate, skin conductance and facial muscle activity were recorded and verbal reports of emotional experience were obtained subsequent to each image. The physiological data, as well as an imagery questionnaire supported the hypothesis that anhedonic subjects were particularly poor imagers and were consistent with data from other laboratories (Lang, 1984) that implicated imagery as a fundamental aspect of emotion processing in humans.
![]()
Anhedonia refers to an inability to experience pleasure. It is generally a transient characteristic (state-like) of depressed patients and a more permanent characteristic (trait-like) of a subgroup of schizophrenic patients (Andreasen & Olsen, 1982 Bernstein & Riedel, 1987 Katsanis, Iacono, Beiser & Lacey, 1992 Rado, 1956). It is frequently assessed with the Physical Anhedonia Scale (Chapman, Chapman & Raulin, 1976), a 61-item true/false inventory that attempts to access a wide variety of positive physical experiences. Most research involving anhedonic subjects has focused on the predictive relationship between anhedonia and future psychopathology (e.g. Chapman, Edell & Chapman, 1981 Haberman, Chapman & Numbers, 1979 Simons, 1981 Simons & Russo, 1987) and very little research has been done which specifically targets emotional responding in anhedonic subjects. Furthermore, since anhedonia is defined as an inability to experience pleasure (i.e. positive emotion), the Physical Anhedonia Scale does not include items to assess experiences of negative affect. Thus, it has not yet been determined whether subjects identified with this instrument have an emotion-processing deficit that is specific to positive emotion, as the definition would suggest, or whether the deficit is more general and includes the entire range of emotional behavior as the clinical descriptor, 'flat' affect, might imply.
There have been two recent attempts to investigate emotional responses of subjects reporting physical anhedonia and they have reached somewhat different conclusions. In the first study, Berenbaum, Snowhite, and Oltmanns (1987) presented film clips, chosen to elicit a variety of emotional experience, to both anhedonic and control subjects. While subjects were viewing each film, their facial expressions were videotaped for off-line coding of emotion. After each film clip, the subjects provided verbal reports of their discrete emotional reactions. No differences were noted between the two groups in either verbal report or their facial expressions.
More recently, Fitzgibbons and Simons (1992) elicited emotional responses from anhedonic and control subjects with color slide stimuli (Lang, Ohman & Vaitl, 1986). Using Lang's 'three-systems' approach to emotion assessment, physiology (heart rate and skin conductance), behavior (facial expression and viewing time), and self report (ratings of valence and arousal) were measured in association with each of the 21 emotion-eliciting slides. Unlike Berenbaum et al. (1987), Fitzgibbons and Simons found evidence for differences between anhedonic and control subjects in all three response systems. In each case, the difference was valence related. Anhedonic subjects rated the slides less positively than controls and this difference in ratings between the two groups increased as the slides became more positive. The valence-sensitive heart-rate response was insensitive to valence among anhedonics at the same time, the anhedonic subjects produced a facial-muscle response that was exaggerated relative to the facial-muscle response of the controls. No differences were noted on any of the measurements specifically related to arousal (i.e. verbal report, skin conductance or slide viewing time).
Though it is difficult to reconcile the findings of Fitzgibbons and Simons (1992) with those of Berenbaum et al. (1987), together the two studies suggest that the emotion-processing differences that exist between anhedonic and normal subjects are both subtle and intriguing. They are subtle since the two experiments shared much in common yet produced divergent results. Subject selection was similar there was overlap in the systems targeted for measurement and the stimulus materials were somewhat similar. Perhaps the main difference between the two studies was one of sensitivity. As suggested by Fitzgibbons and Simons, when facial expressions are coded from videotaped records as they were by Berenbaum et al., actual movement of the muscles is required before expressions can be detected and scored. Facial EMG, however, can detect 'covert expressions'-changes in the same muscles, but not of sufficient magnitude to produce actual changes in facial features. Likewise, it may be that the assessment of self-reported valence across a wide variety slide content may be more sensitive in the verbal-report domain than requiring subjects to discriminate and judge among multiple discrete emotion categories as they did in Berenbaum et al. Heart rate, sensitive to both valence and subject group in Fitzgibbons and Simons, did not have a parallel in the Berenbaum et al., experiment.
The between-group differences are intriguing for a variety of reasons. In particular, they suggest cognitive mechanisms which may underlie the differences and thereby point to further experimentation. As indicated above, the measures employed by Fitzgibbons and Simons-i.e. self report, covert facial expression, and heart rate, correspond to the language, behavioral, and physiological systems discussed by Lang in his three-systems view of emotion (e.g. Lang, 1984 1985). In fact, the three-system profile obtained from the anhedonic subjects seems to exemplify the kind of systems-desynchrony that gave rise to Lang's three-systems approach initially. Anhedonic subjects failed to produce an integrated or synchronized pattern of affective response. If one considers the aggregate control-subject response to exemplify what Lang might call the positive-emotion prototype, it may be that this prototype is either absent or not well integrated (incoherent) in anhedonic subjects. This may not be the case for the negatively-valent emotions.
Another implication of the Fitzgibbons and Simons data is that anhedonics may be a group of subjects with poor imagery ability. This conclusion stems from Lang's argument that changes in visceral physiology indicate that subjects have accessed the deep structure of the emotion while verbal report, and possibly facial expression, can be heavily influenced by the semantic information in the emotion network and may reflect only an appraisal of the stimulus content. Lang suggests that the deep structure of the emotion contains the motor program and the physiological support for an organized emotional response. Activation of this structure, which Lang refers to as 'image-mode' processing, necessarily involves measurable outflow. Since Lang (1984) has shown in several studies that subjects with poor imagery ability fail to produce an appropriate visceral physiology to emotional stimuli, it may be that the insensitivity of heart rate to the valence of the slide stimuli noted by Fitzgibbons and Simons in their anhedonic subjects indicates that one aspect of anhedonia is poor imagery ability.
The present study was designed to investigate imagery ability in subjects with physical anhedonia and at the same time probe the issue of emotion-prototype integration by including imagery scripts targeted at a variety of both positive and negative emotions. Imagery ability was inferred both by the degree of visceral activation which occurred during the imagery task and with the Questionnaire on Mental Imagery (QMI Sheehan, 1967). Activity from both the zygomatic and corrugator muscles was recorded along with heart rate and skin conductance. Self-report of emotion experience was collected in a discrete-emotion format using Zuckerman's Inventory of Personal Emotional Reactions (ZIPERS: Zuckerman, 1977), and measures of valence and arousal were derived from the ZIPERS through principal components factor analysis. It was hypothesized that anhedonic subjects would report poorer imagery ability on the QMI and that this deficit would be associated with reduced visceral activation during the imagery task. It was expected that group differences would be associated with the valence aspect of emotion and be most apparent when processing positive imagery scripts.
An additional purpose of the present research was to establish a psychophysiological link between the dimensional approach to emotion assessment and research and the discrete-emotion approach. As indicated above, previous research has demonstrated that facial EMG and heart rate are specifically related to emotion valence while skin conductance is related specifically to arousal. In all cases in which these relationships have been demonstrated, however, the valence and arousal ratings have been obtained directly with Lang's Self-Assessment Manikin (SAM Lang, 1980). Since these dimensions were originally derived from the language of discrete emotions (Mehrabian & Russell, 1974 Russell & Mehrabian, 1977), it was a specific aim of the present experiment to employ the discrete-emotion language, attempt to derive measures of emotional valence and arousal, and establish whether the specific relationships between these dimensions and the psychophysiological measures described in previous research would hold under the present conditions as well.
Sixty-four undergraduate students took part in this study and received partial credit toward the research participation component of their introductory psychology course. Students who scored two standard deviations above the mean on the Physical Anhedonia Scale (n=33 17 female) were identified as anhedonic while control subjects (n=31, 16 female) were selected randomly from among those students whose questionnaire scores were less than +0.5 standard deviations above the mean. The experimenter (EF) was blind to all subjects' group membership.
Eight scenes were developed, based on interviews with undergraduate students, which would reliably elicit a range of discrete emotional reactions. In addition to this standard 8-item imagery set, instructions for two additional scenes were provided which cued subjects to image both an idiosyncratic negative and positive 'prototype'-events from their personal lives which when revisited would elicit strong affective responses. The specific content of these two images remained private. Instructions for all scenes were tape recorded and constituted the stimuli used in the present experiment. The imagery scripts, along with their target emotions, are contained in Table 1.
---------- Table 1 ----------
Self-report. The Zuckerman Inventory of Personal Emotional Reactions (ZIPERS) is a thirteen item scale designed to assess specific emotions such as fear, anger, happiness etc., and two items to obtain self reports of visceral arousal (heart rate and respiration). Each item (e.g. "I feel afraid") requires subjects to rate the intensity of their emotional reaction on a five-point scale. The complete inventory is contained in Table 2.
---------- Table 2 ----------
Physiological Measures. An electrocardiogram (EKG) was recorded by placing Med Associates Ag-AgCl miniature electrodes over the subjects' left and right anterolateral rib cage. The EKG signal was amplified by a Coulbourn Model S75-01 Hi-Gain Bioamplifier and fed into a Coulbourn Model S21-06 Bipolar Comparator which produced a digital output upon the detection of each 'R'-wave in the EKG.
Skin conductance was measured using a Coulbourn Model S21-22 constant voltage (0.5V) skin conductance coupler. Thenar and hypothenar electrode sites on the non-dominant palm were cleansed with distilled water and standard Ag-AgCl electrodes (0.5 cm2) were applied. Johnson and Johnson K-Y Jelly served as the electrolyte.
Bipolar electromyographic (EMG) recordings were made from Med-Associates miniature Ag-AgCl electrodes placed along the zygomatic and corrugator muscles. The EMG signals were amplified with Coulbourn High Gain Bioamplifiers (Model S75-01 bandpass 8 to 250 Hz) and the output was passed to Coulbourn Cumulating Resetting Integrators (Model S76-22) for full-wave rectification and integration over time.
During the experiment, the tape-recorded standardized scenes and instructions to image the two personally constructed scenes were presented to each subject over headphones. The description of each scene lasted 30 seconds and was followed by a 20 second imagery period and a 20 second recovery period. During the imagery period, subjects were instructed to imagine the scene that had just been presented as vividly as possible, i.e. as if they were actually participating in the activity described. The format of the two idiosyncratic (nonscripted) imagery trials was similar. On the ninth trial, subjects were instructed during the presentation period to imagine the event from their own life experience that they recalled as particularly negative, and on the final trial they were instructed to recall the life event that they had chosen as particularly positive.
At the outset of the experiment, all subjects were required to complete the imagery questionnaire (QMI) and at the conclusion of each imagery period, subjects were asked to complete their ZIPERS ratings using an Apple IIe computer.
Heart rate, skin conductance and integrated facial EMG were recorded on each trial prior to the delivery of the imagery script, during delivery, and during the visualization period. The data were recorded on-line using a Digital Equipment Corporation PDP-11/34 laboratory computer that was synchronized with the tape recorder containing the imagery scripts. Heart rate was simply quantified by calculating the mean in beats per minute for each period. Analog skin conductance was sampled at 42 cps and then displayed off-line on a graphics terminal where each response that exceeded .06 microSiemens (uS) was visually identified and tallied. The number of resets from the integrators of zygomatic and corrugator EMG were counted during each period and then transformed into a single EMG pattern score by subtracting the number of corrugator from zygomatic integrator resets. This derived score served as the EMG dependent variable (Greenwald, Cook & Lang, 1989). A positive pattern score reflected a relative increase in zygomatic activity and a negative pattern score was indicative of a relative increase in corrugator activity. For each of the three physiological measures, scores for the presentation and visualization periods were computed as change from the predelivery base period mean.
![]()
As a first test of the hypothesis that anhedonics were similar to poor imagers, the QMI data from the two groups were compared. The QMI is a 35 item scale which requires subjects to rate the vividness of their imagery. High scores on the QMI are associated with poor imagery. In support of the poor imagery hypothesis, anhedonic subjects scored higher than controls (91.7 v 79.2). This difference was significant (t(62)=1.88, p<.05, one-tailed).
The analysis of the emotion self-report was based on the factor analysis of the entire set of ZIPERS data. The factor analysis revealed two factors with eigenvalues greater than 1.0 and these two factors accounted for 70% of the data-space variance. The factor loadings are presented in Table 3. As the table indicates, the first, and most substantial factor, grouped all the positive-emotion questions at one pole and all the negative-emotion questions at the opposite pole. This was tentatively identified as the valence factor. The second factor was composed essentially of the two physiology items. This factor was identified with arousal.
---------- Table 3 ----------
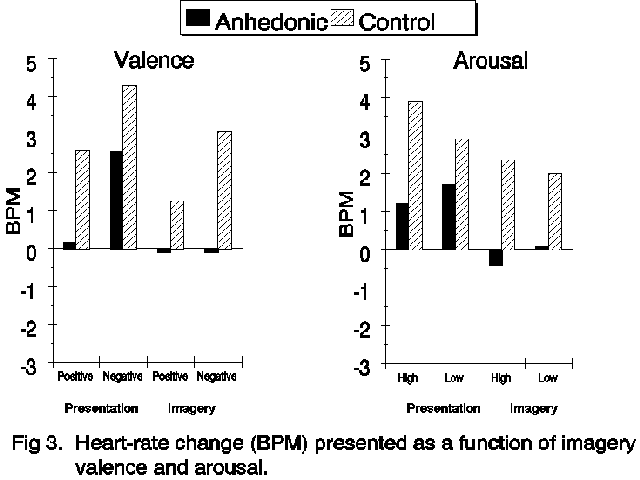
As an initial examination of the differences between emotion self-report in anhedonic and control subjects, a Group (2) X Scene (2) mixed-model ANOVA was conducted on both valence and arousal scores by first grouping the five most positive scenes together and comparing them to the five most negative scenes and then by grouping and comparing the five most arousing to the five least arousing scenes. Both valence and arousal data are presented in Figure 1. The figure indicates that substantial group differences occurred
---------- Figure 1 ----------
on both emotion dimensions. Anhedonic subjects rated the scenes overall as less pleasant (F(1,60)=17.90, p<.001), though a significant interaction between subject group and valence was also present (F(1,60)=18.00, p<.001). Bonferoni adjusted 't'-tests indicated that the between-group differences were limited to scenes with positive valence. As Figure 1 indicates, the 'positive' scenes were rated as significantly less positive by anhedonic than control subjects while there were no significant differences between the two groups on the 'negative' scenes.
Anhedonic subjects also rated the imagery scenes as less arousing than control subjects did (F(1,60)=12.30, p<.001). This difference between groups held for both positive and negative scenes, though there was a significant interaction between Group and Arousal category (F(1,60)=8.60, p<.01) implying that the group difference was greatest when scenes had high arousal value.
As is often true (Fitzgibbons & Simons, 1992 Greenwald et al., 1989), scenes with negative valence were reported to be more arousing than scenes with positive valence (F(1,60)=29.42, p<.001). This was true for both anhedonic and control subjects.
To assess the relationships among valence, arousal and each of the physiological measures, scenes were again divided into valence and arousal categories. Separate ANOVAs were then conducted on each physiological measure during the presentation and imagery portions of each trial.
Facial EMG. Figure 2 depicts the relationship during stimulus presentation and imagery periods between the EMG pattern score and both valence and arousal.
---------- Figure 2 ----------
Facial EMG was strongly related to the valence of the image, both during presentation and visualization (F(1,59)=47.82, p<.001), though an interaction between Period and Valence (F(1,59)=18.47, p<.001) was present and indicates that the effect of valence on facial EMG is larger during visualization than during scene presentation. As seen in Figure 2, positive scenes prompted a relative increase in zygomatic activity while the negative scenes were associated with a more dominant corrugator pattern. This relationship between valence and EMG pattern score was evident in both anhedonic and control subjects.
Among control subjects, there was a weaker, but still significant relationship between facial EMG and arousal with high-arousal scenes associated with a more zygomatic-dominant facial pattern than low-arousal scenes. Statistically, this effect is seen in the interaction of Group and Arousal (F(1,59)=8.42, p<.01). A closer inspection of the data from individual trials (see below) revealed that this unanticipated relationship between arousal and facial EMG was due primarily to a very large zygomatic response during Scene 10 -- the individual-positive scene rated by controls as very positive and very arousing.
Heart rate. The heart rate data are presented in Figure 3, and as the figure illustrates, heart rate responsivity was related specifically to image valence (F(1,58)=4.70, p<.05), not arousal (F<1). There was more HR
---------- Figure 3 ----------
acceleration during scene presentation than during imagery (F(1,58)=16.05, p<.001), and negative content was associated with greater HR acceleration than positive content during both periods. Unlike the EMG data, however, the HR response differed significantly in the two subject groups. Control subjects produced more HR acceleration overall (F(1,58)=9.56, p<.01), and showed equivalent valence effects during both presentation and imagery. Anhedonic subjects were unable to maintain a differential heart rate response during imagery. In fact, during imagery anhedonic subjects were unable to mount any heart-rate response at all. When directional 't'-tests were conducted on the response of each group during positive and negative imagery, both responses from control subjects differed significantly from zero (positive t(31)=1.71, p<.05 negative t(31)=3.19, p<.01) while this was true of neither response from the anhedonic subjects (i.e. both means were below zero).
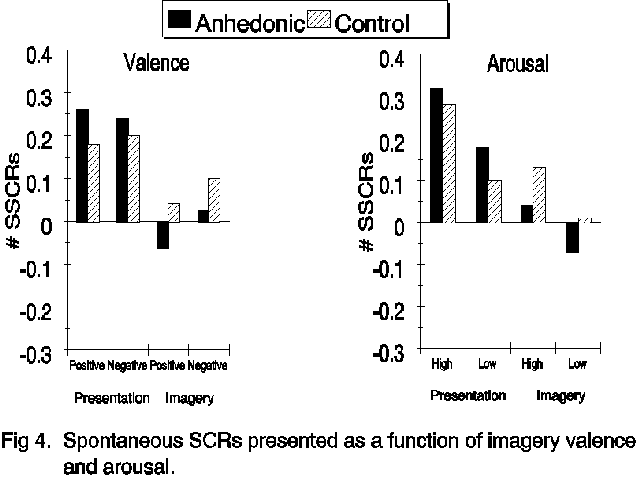
Skin conductance. Spontaneous skin conductance activity is presented in Figure 4. Unlike the muscles of the face and the heart-rate response, there was no significant relationship between the number of spontaneous skin conductance responses (SSCRs) and image valence (F<1). There was, however, a highly significant relationship between skin conductance and arousal during scene presentation and imagery (F(1,62)=12.89, p<.001). As can be seen in Figure 4,
---------- Figure 4 ----------
high arousal scenes were associated with more frequent spontaneous skin conductance responses and this relationship was equivalent in the two subject groups (Group X Arousal F<1). Responses were less frequent during imagery than during presentation (F(1,62)=24.84, p<.001). This was true in both groups and there was no evidence that anhedonic and control subjects differed in their overall production of spontaneous SCRs (F<1).
A more fine-grained analysis of the present data was conducted by examining the imagery periods from the five negative and five positive emotions independently for evidence of physiological specificity and to assess the possibility that between-group differences might be specific to one or more discrete emotions.
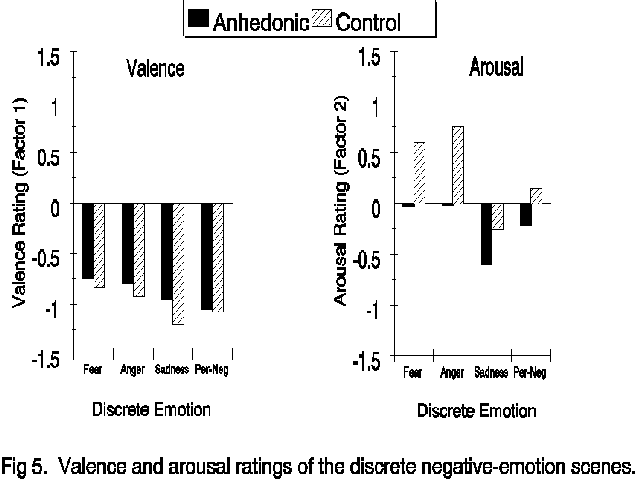
Discrete negative emotions. To assess the possibility that differences might be found among the negative emotions, the self-report and physiological data from the two fear scenes were combined and compared to the data from the anger, sadness and personal-negative scene using Group X Emotion Mixed-Model ANOVAs. Figure 5 contains the Valence and Arousal ratings for the negative scenes. Significant
---------- Figure 5 ----------
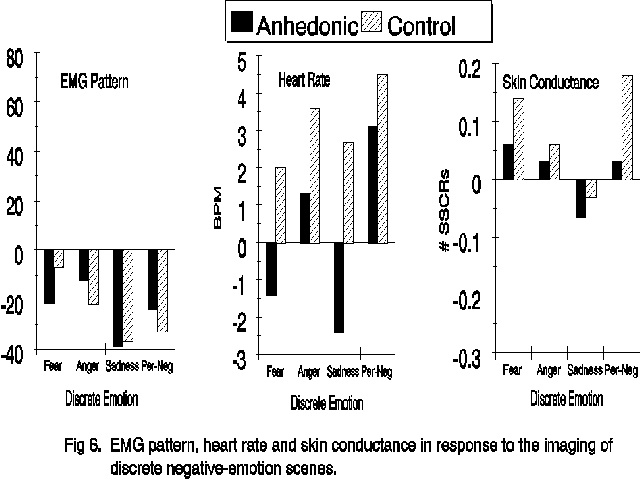
self-report differences among the imagery scripts were apparent on both the Valence (F(3,180)=29.03, p<.001) and Arousal (F(3,180)=22.40, p<.001) factors of the ZIPERS. Post-hoc Newman-Keuls tests indicated that the sadness and the individual-negative scripts evoked images that were rated as more negative and less arousing than the fear or the anger scripts. Significant Group (F(1,60)=4.89, p<.05) and Group X Script (F(3,160)=3.14, p<.05) valence effects indicated that anhedonic subjects rated the fear, sadness and anger scripts less negatively than did the control subjects while the personal-negative imagery was rated as equally unpleasant by the two groups. A similar pattern was observed in the subject reports of arousal, though only the main effect of Group membership reached statistical significance (F(1,60)=7.00, p<.05). The results of the analyses on the physiological data revealed no difference among the negative-emotion scenes and save for the reduced heart-rate response from anhedonic subjects already described (F(1,58)=7.82, p<.01) , there were no significant physiological effects involving group membership when only the negative emotions were considered. These data are presented in Figure 6.
---------- Figure 6 ----------
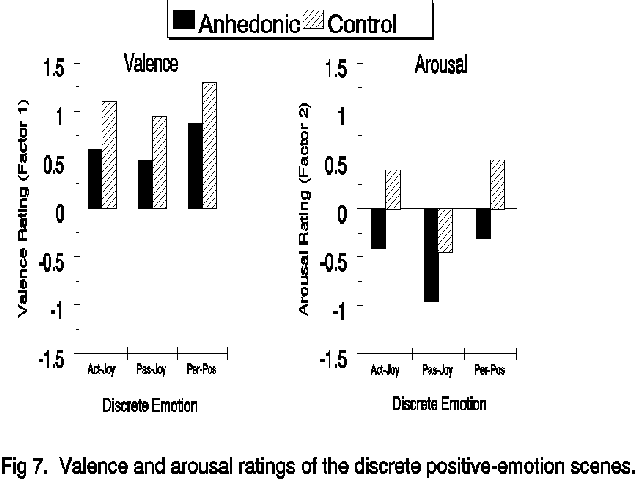
Discrete positive emotions. To analyze the positive emotions, Scripts 1 and 5 were combined and defined as Active Joy and and Scripts 3 and 6 were combined and defined as Passive Joy based on the distinction made by Izard (1991). The self-report and physiological data from the associated imagery along with the data from the personal-positive scene were submitted to analysis of variance as described above. The self-report data are presented in Figure 7. The ANOVAs and post-hoc Newman-Keuls tests revealed that subjects rated the active joy and personal-positive imagery as more positive (F(2,120)=41.99, p<.001)
---------- Figure 7 ----------
and more arousing (F(2,120)=51.35, p<.001) than the passive joy imagery. The personal-positive image was rated higher on valence than either active and passive joy, though the personal-positive and active-joy images were equally arousing. The pattern of self-report ratings was similar in the two groups (i.e. there were no Group X Script interactions), though anhedonic subjects rated all positive images as less positive (F(1,60)=20.84, p<.001) and less arousing (F(1,60)=15.40, p<.001) than did the control subjects.
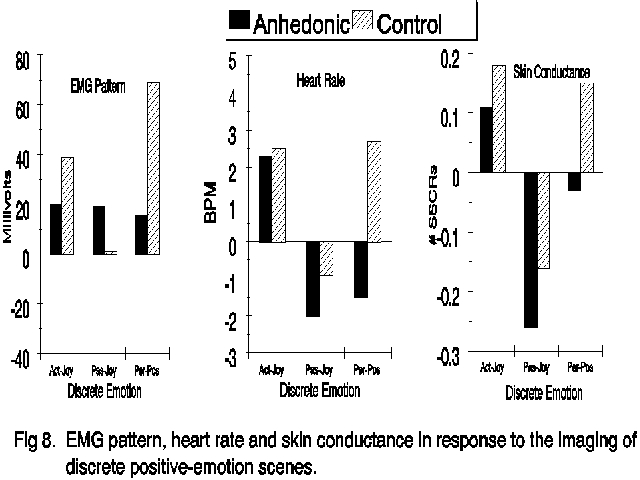
The physiological responses were closely related to the distinction among the positive emotions. The data are contained in Figure 8. Consistent with the arousal difference between active and passive joy, there were also
---------- Figure 8 ----------
significant differences between positive emotions on the frequency of nonspecific skin conductance fluctuations (F(2,124)=8.22, p<.001). Both active joy and the personal-positive image prompted greater SC activity than did imagery of passive joy, and this was true in both subject groups. Likewise, the two valence-sensitive physiological measures also distinguished among the positive emotions. Significant main effects occurred for heart rate (F(2,116)=4.19, p<.05) and facial EMG (F(2,118)=4.87, P<.01). For control subjects, active joy and the personal-positive emotion were characterized by very similar heart rate and facial EMG responses, and each exceeded the response which occurred during passive joy. Anhedonic subjects, on the other hand, produced heart-rate and facial EMG responses during personal-positive imagery that more closely resembled their response during passive joy than during active joy. The interaction between Group and Emotion was statistically significant for EMG (F(2,118)=6.21, p<.01) but it was not for heart rate (p>.25). Exploratory ANOVAs conducted separately on the HR and EMG data from the two groups revealed significant differences in the control group among the positive emotions for for both facial EMG (F(2,64)=11.52, p<.01) and HR (F(2,62)=3.15, p<.05) but no difference on either measure was found among the positive emotions in the anhedonic group (p>.10).
![]()
One purpose of the present experiment was to explore the generalizability of the dimensional approach to human emotion research. The experiment was very successful in that regard. Though the stimuli employed in this experiment were developed to elicit discrete emotions and the self-report instrument (ZIPERS) was designed to assess emotion experience from a discrete-emotion perspective, the present data actually provide good support for the utility of the dimensionality approach to emotion research (Russell & Mehrabian, 1977 Lang, 1984 Lang, 1985). This was particularly evident in the relationships between the physiological measures and the self-report factors obtained from the ZIPERS ratings. Lang and his colleagues (e.g. Greenwald et al., 1989), along with Fitzgibbons and Simons (1992), have shown that facial EMG and HR relate to emotion valence and SCR relates to arousal when valence and arousal are assessed directly. These same relationships were evident in the present data when valence and arousal were defined through linear combination of discrete-emotion ratings. Facial EMG and HR were strongly related to Factor 1 (Valence) and weakly or unrelated to Factor 2 (Arousal) the opposite was true for fluctuations in skin conductance. This is particularly interesting since not only verbal report but also visceral physiology (HR and SSCR) was assessed differently here than it was by both Greenwald et al. (1989) and by Fitzgibbons and Simons (1992). For example, rather than assessing the acceleratory peak in a phasic heart rate response, the present study used the more tonic measure of mean heart rate. While during imagery, high heart rate was associated with negative valence and not positive valence as it was when phasic responses to discrete stimuli are assessed, it was still valence and not arousal that determined the response. Similarly, in the present study skin conductance was assessed by counting spontaneous fluctuations, not by measuring the amplitude of discrete responses. Still, the specific relationship of skin conductance to arousal remained unequivocal. Given the substantial variation across studies and laboratories in both stimulus and response methodology, it would appear that the relationships among these physiological response systems and the emotion dimensions are strikingly robust.
The utility of the dimensional approach was particularly evident when valence and arousal were considered together in the classification of the emotion scripts. As evident in Figures 5 through 8, there were clear differences among the scripts within both positive and negative valence categories and these differences could be accounted for by each script's position along the arousal dimension. Within the negative-emotion scenes, those designed to elicit fear and anger were rated as more arousing than the scene eliciting sadness, and within the group of positive-emotion scenes, the two active-joy scenes were more arousing than the two passive-joy scenes. It is important to note that while the verbal report of arousal (ZIPERS Factor 2) was initially used to subtype the scenes within each valence category, the identical subtypes would have resulted if the subtyping had been done on the basis of the skin conductance recordings. This specificity in the relationship between physiology and the two emotion dimensions was compelling and argues strongly for the utility of the dimensional approach to emotion.
Another aim of the present experiment was to determine whether anhedonic and control subjects differed in imagery ability. Specifically, it was hypothesized that anhedonic subjects would be characterized as poor imagers. The data supported this hypothesis. In the paper and pencil domain (i.e. the QMI) , anhedonics scored significantly higher than control subjects, with high scores indicative of poor imagery. In the physiological domain, the heart-rate of the anhedonic subjects seemed to be dissociated from the imagery task, particularly during unassisted visualization. This is important since Lang (see Lang, 1984) has shown that the heart-rate response reliably distinguishes between good and poor imagers. In Lang's work, poor imagers do not seem to engage the heart-rate response system when generating emotional imagery and the same failure to produce a consistent heart-rate response was noted in the anhedonic subjects who served in the present experiment.
The primary purpose of the present study was to evaluate the three-system emotional response in anhedonic and control subjects in order to shed further light on the nature of the emotion-processing deficit associated with anhedonia. Though there were some differences between the present data and those described by Fitzgibbons and Simons (1992), evidence of an emotion-processing deficit was again observed in subjects reporting physical anhedonia.
Clear differences between anhedonic and control subjects were observed in their verbal report of emotion experience. In Fitzgibbons and Simons (1992), this difference was restricted to ratings of emotional valence, but in the present study, this difference was evident in ratings of both valence and arousal. Between-group differences were also noted in the heart-rate response to affective stimuli. As was true in Fitzgibbons and Simons (1992), heart-rate and valence were significantly related in control subjects but this relationship was absent in subjects with anhedonia. Thus, two of the three response systems (verbal report and visceral physiology) involved in emotion production were sensitive to group membership.
The only measure of overt behavior included in the present study was facial EMG. As expected, it was clearly a valence-related measure, but a measure only weakly related to group membership. Unlike Fitzgibbons and Simons, who found that anhedonics produced more facial EMG than did control subjects, the only facial-EMG difference between the two groups noted in the present study (Figure 8) was a reduced, not enhanced, covert expression from anhedonic subjects when imagining their personal-positive scene. Though facial expression is far from a perfect index of emotion-related overt behavior (see Lang, 1985), we have also on two different occasions (see Simons et al., in press), surreptitiously measured the length of time subjects chose to view individual color-slide stimuli that varied on valence and arousal. Though there was a tendency on both occasions for anhedonics to view slides for a shorter period of time than control subjects viewed them, these differences were not reliable on either occasion. Thus, while anhedonic and control subjects consistently differ in the verbal report and visceral components of emotion, laboratory evidence of overt-behavioral deficits has been much more difficult to obtain.
Based on the pattern of differences between anhedonic and control subjects that emerged in response to emotion-eliciting color-slide stimuli, Fitzgibbons and Simons (1992) concluded that the emotion-processing deficit that characterized the anhedonic subjects extended beyond verbal report, was valence related and was most likely specific to the positive pole. The present data, however, suggest that the differences between groups may be more complex than that. First, during imagery, between-group verbal report differences were present not only as a function of image valence but as a function of arousal as well. Second, the verbal-report differences were present in response to both positively-valent and negatively-valent images. Third, during imagery, anhedonic subjects produced no consistent HR response to either positive or negative imagery scripts. Thus, the present data, though still highlighting positive emotion, suggest an emotion-processing deficit that may run through the entire valence dimension and may involve arousal mechanisms as well. In fact, the 'discrete-emotion' data (see Figures 7 & 8) suggest that a complex interaction between the two emotion dimensions and the imagery process may best characterize the deficit.
Recall that when control subjects were instructed to imagine a situation from their past in which they felt wonderful and thought things were just perfect, they generated an emotion pattern resembling active joy. Anhedonic subjects, given the same instructions, generated a passive-joy pattern at best. With color-slide stimuli (Fitzgibbons & Simons, 1992), or with very specific imagery scripts (e.g., #s 1 & 5), anhedonic subjects can, more or less, produce an active-joy emotion. When no auxiliary cues are provided (e.g., #10), they fail.
As mentioned above and detailed elsewhere (Simons, Fitzgibbons and Fiorito, in press), the basic difference between anhedonic and normal control subjects may involve the way in which emotions are structured in long-term memory. Lang (1985) has suggested that memorial representations of emotion may vary in coherence. A coherent representation is one in which associations are strong and one which can be accessed in toto even with highly degraded stimuli. Incoherent representations are only loosely structured and require a more direct match with the invoking stimulus before complete processing of the emotion will occur.
The present data raise the possibility that in anhedonic subjects there is relative incoherence in the structure of fundamental emotional representations and that poor coherence may be particularly characteristic of positive-emotion representation. With incoherent memorial representations, it is difficult to provoke 'image-mode' processing and to observe its associated visceral outflow. As indicated above, this difficulty would increase as stimuli become increasingly degraded (i.e., from real to imaginal). It is suggested that this profile of less coherence overall, and less coherence of positive- than negative-emotion 'prototypes' in the long-term memory of anhedonic subjects would explain the pattern of between-group differences noted here and in our previous study with more concrete stimuli (Fitzgibbons and Simons, 1992).
Fitzgibbons and Simons (1992) noted only valence-related deficits in their study and suggested that the memorial representation (prototype) that organizes the positive-emotion response may be absent or incoherent among anhedonic subjects (see Lang, 1985).
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}