* Address reprint requests to W.M. Perlstein, Department of Clinical and Health Psychology, University of Florida, P.O. Box 100165 HSC, Gainesville, FL 32610. E-mail: wmp@grove.ufl.edu
This research was conducted in partial fulfillment of the Ph.D. dissertation by the first author and was supported by MH42465 from the National Institute of Mental Health to Frances K. Graham and Robert F. Simons.
Portions of the work were reported at the 34th Annual Meeting of the Society for Psychophysiological Research in Atlanta, October 1994.
![]()
A
putative gating mechanism reduces startle blink, midline scalp potentials
beginning with P50, and perceived loudness of startling stimuli.
Tactile prestimuli were paired with auditory startle stimuli to determine
if: 1) P50 inhibition is due to an extrinsic mechanism, 2) pairing
differentially affects potentials reflecting modality specific and nonspecific
system activity, and 3) crossmodal pairing modifies perceptual magnitudes of
both pair members. Stimuli were
presented alone and in pairs separated by 60 or 360 ms.
ERPs were recorded from midline and lateral sites; EMG was recorded from
several facial and scalp muscles. Pairing
reduced blink, and midline P50, N100 and P200 amplitudes; reductions were
greater at the longer interval. P30
was largely unaffected by pairing. Pairing
also differentially affected lateral N100 components reflecting later activity
in specific and nonspecific systems. Results
show that prestimulus inhibition of ERPs is not due to intrinsic refractoriness
and that pairing differentially affects ERPs associated with modality specific
and nonspecific projection systems.
![]()
Prepulse inhibition (PPI)—a reduction in the amplitude of the startle reflex that occurs when a weak stimulus (S1) shortly precedes a reflex-eliciting stimulus (S2)—has been proposed to index an automatic “gating” mechanism that protects incoming information from being assimilated with or distorted by a shortly-following competing input (Graham, 1975). The concept of a gating mechanism implies an extrinsic neural mechanism rather than intrinsic refractoriness in a common input path. Blink PPI, for example, is widely believed to reflect the activity of an extrinsic mechanism mediated via a parallel indirect path, external to but synapsing with the direct path that underlies the unmodulated startle response (Davis, 1984; Fendt & Koch, 1999; Koch & Schnitzler, 1997; Kodsi & Swerdlow, 1997a, b; Leitner, Powers, & Hoffman, 1979; Leitner, Powers, Stitt, & Hoffman, 1981; Saitoh, Tilson, Shaw, & Dyer, 1987; cf. Schicatano, Peshori, Gopalaswamy, Sahay & Evinger, 2000). In addition to the anatomical data, findings indicating attentional modulation of blink PPI point to the involvement of an extrinsic mechanism (e.g., Filion, Dawson & Schell, 1993; Hackley & Graham, 1987). However, if such a gating mechanism is to account for perceptual effects, it should not be limited to effects on startle blinks, but should also produce concomitant changes in exogenous or obligatory sensory components of the scalp-recorded event-related potentials (ERP) as well as perception. Perlstein, Fiorito, Simons, and Graham (1993) confirmed that, compared to S2s presented alone, acoustic S1s preceding acoustic S2s reduced reflex blinks and S2-elicited ERP midline components transmitted via the modality nonspecific (i.e., extralemniscal) projection system (i.e., P50, N100); no pairing effects were seen on the earlier P30 component, believed to be associated with activity in the modality specific (i.e., lemniscal) projection system. Further, judged loudness decreased for S2 and increased for S1, indicating some degree of perceptual assimilation.
The present research was designed to replicate and extend the findings of Perlstein et al. (1993) in order to: 1) exclude refractoriness as an explanation of prepulse inhibition effects on ERPs; 2) evaluate prepulse effects on laterally-recorded later-latency ERPs that similarly reflect activity in the specific and nonspecific sensory projection systems; and 3) further evaluate perceptual concomitants of prepulse inhibition.
First, to exclude the possibility of refractory effects of S1 on the response to a same-modality S2 in a common (i.e., same modality) input path, we used crossmodal tactile-acoustic pairs and appropriately varied the intrapair interval. The crossmodal pairings employed were selected to yield blink PPI with lesser inhibition at a short than long stimulus onset asynchrony (SOA) and, because different-modality stimuli activate different afferent paths, to eliminate confounding modality-specific repetition or refractory effects that occur with intramodal pairs for both blink (Balaban, Anthony, & Graham, 1985; Rimpel, Geyer, & Hopf, 1982) and ERPs (Davis, Osterhammel, Wier, & Gjerdingen, 1972). Notably, the temporal course of blink PPI differs from the course of refractoriness. Refractory effects peak at near simultaneity of two stimuli and decay monotonically thereafter but prepulse inhibition reaches its maximum after a delay. For acoustic S2s, blink inhibition has been found to peak at SOAs of about 120 ms when paired with acoustic S1s and about 250 ms with tactile and visual S1s; the initial 25- to 50-ms period following tactile or visual prestimuli may even enhance the blink elicited by an acoustic S2 (Blumenthal & Gescheider, 1987; Blumenthal & Tolomeo, 1989; Graham, 1980).
To demonstrate temporally-delayed inhibition, therefore, it is necessary to show that inhibition is less at some short than longer SOA. The short SOA must be long enough that two stimuli are individually perceived, but not close to or beyond the point of maximal inhibition. If the short SOA is near or beyond the point of maximal inhibition, response magnitude at any longer SOA would reflect a monotonically increasing function. Although Perlstein et al. (1993) demonstrated acoustic-pair inhibitory effects in two experiments, they could not exclude refractoriness on the basis of a delayed maximum inhibition because the SOA in the initial study was 120 ms (i.e. at the point yielding maximal blink inhibition in previous studies) and, in the second study of another group of subjects, the SOA was 500-ms. The longer interval is close to the upper limit for obtaining any inhibition. Consequently, comparing short with long SOA effects under these conditions might reflect only the recovery phase of a refractory process and would not be adequate to differentiate intrinsic and extrinsic effects To avoid measuring recovery and refractory effects in the present research, we selected, based on pilot data with tactile-acoustic pairs, a short SOA of 60 ms and a long SOA of 360 ms. Although blinks are enhanced at lead intervals shorter than 60 ms and are inhibited at intervals between 60 and 360 ms, we chose a 60-ms SOA as the short interval to achieve good temporal separation between S1 and S2 and a 360 ms SOA as the long interval to avoid near-maximal inhibition.
Second, to examine pairing effects on later-latency ERP components believed to be generated in auditory cortex, we recorded from two lateral leads in addition to the two midline leads used by Perlstein et al (1993). The Perlstein et al. study identified five components: P30, N40, P50, N100 and P200. Of these, P30 was not inhibited by pairing, N40 was enhanced, and beginning with P50, the remaining components showed significant reductions. The literature suggests that P30 and P50 not only arise from different thalamo-cortical generators, but also reflect activity in different regions of auditory cortex—primary and secondary or association areas (Celesia, 1976; Scherg & von Cramon, 1986), respectively—and are activated in parallel by different neural projection systems. Specifically, P30 may reflect activity transmitted via a rapidly-conducting, modality-specific path which initiates the transient-dependent prepulse effects in a more rostral and medial, slowly-conducting, polysensory path leading to the P50 generator. Consonant with a specific system, P30 recovers rapidly from stimulation, remains stable at rates as fast as 10/sec, and is unchanged across different sleep states. In contrast, P50 is markedly reduced or absent at stimulation rates of 5/sec and, although unchanged during the awake and REM-sleep states, is depressed during slow-wave sleep. Further, P50 is more sensitive than P30 to barbituate anesthetic agents and, unlike P30, is diminished by cholinergic blockade (Dickerson & Buchwald, 1991; Deiber, Ibañez, Bastuji, Fischer & Mauguiére, 1989; Erwin & Buchwald, 1986a, 1986b, 1987; Harrison, Woolf, & Buchwald, 1990).
In contrast to P30 and P50, N100 and P200 may reflect activity of both specific and nonspecific systems. Little is known about the sources of P200, but Näätänen and Picton (1987), based on a comprehensive review, described three components of N100 that have different functional characteristics and can be independently assessed. Component 3 (C3) has characteristics of a nonspecific response system and is the major contributor to the midline N100. It is elicited by auditory, visual, and tactile stimuli, has a recovery time of at least 10 s, and increases with sound intensity up to 90-100 dB. Component 1 (C1) has a source in or near primary auditory cortex and shows characteristics of a specific system—its recovery period is short and it increases with intensity only up to 60-80 dB. Component 2 (C2) can be identified over temporal regions as a biphasic “T-complex” consisting of a small P105 (Ta) and a larger, later negativity (Tb or C2) that may lie within auditory association cortex. Here, we refer to the Tb deflection as Component 2. It appears to share some of the functional characteristics of both the specific and nonspecific components. Therefore, the present research, in addition to assessing prepulse effects on ERPs at midline sites, also assessed effects on specific-system N100 activity (i.e., Components 1 and 2) at two lateral leads.
Third, the present research determined whether magnitude estimates of different modality pairs would replicate our earlier finding of increased perceptual magnitude of S1 and decreased perceptual magnitude of S2 compared to magnitude estimates of either stimulus presented alone. Filion, Dawson, and Schell (1998) have recently reviewed studies, using other perceptual measures that also provide some support for the prepulse “protection-of-processing” hypothesis.
![]()
Participants were 36 undergraduate students (15 female); seven were paid $10 for their participation, all others received course credit in introductory psychology. They were given a description of procedures and completed a medical screening questionnaire prior to providing written informed consent. Three subjects were excluded and replaced because the eye-movement correction procedure (see below) rejected more than 30% of ERP trials in at least one condition. All subjects met the minimum criterion of at least 50% acceptable non-zero blink trials in the single, control-tone condition.
An 800-Hz, 110-dB SPL(A) tone duplicated the S2 of Perlstein et al. (1993) and a 10-psi airpuff psychophysically matched their 75-dB S1 tone; standard stimuli, anchoring magnitude estimates, were psychophysically-matched at 22-psi and 90-dB. All stimuli had 40-ms durations and near-instantaneous rise time. Puffs were gated by computer-controlled solenoid stopcocks and delivered to the subject through Tygon tubing terminating in a short piece of pliable lead tubing mounted in foam rubber. The 2-mm diameter nozzle of the lead tube was positioned on the right forearm between tendons of palmaris longus and flexor carpi radialis muscles, 2-cm proximal to the wrist crease, the upper-extremity site commonly used to elicit somatosensory potentials electrically (Chatrian et al, 1984). The nozzle was affixed to the subject with ACE bandaging and the delivery tubing was concealed by a blanket wrapped around the arm, effectively eliminating any acoustic artifact1.
Acoustic stimuli of 75, 90, or 110 dB, generated by a Coulbourn Precision
Signal Generator (S81-06), were gated through a Coulbourn Programmable
Attenuator (S85-08) and, with continuous 60-dB white noise from a Coulbourn
Noise Generator (S81-02), were fed through a Coulbourn Audio-Mixer Amplifier
(S82-24) for binaural delivery via TDH-49P earphones mounted in a Tracor RA-125
headset.
Physiological
activity was recorded from 10 leads: 2 EOG and 4 EMG (impedance < 10kohms); 4
electroencephalographic (EEG; impedance < 5 kohms).
Electrodes for vertical and horizontal EOG were centered 13-mm above and
below the right eye and on the outer canthus of each eye.
Bipolar EMG, measuring blink, was recorded from laboratory-fabricated
electrodes (2-3 mm diameter; Silverstein & Graham, 1978) centered 3-mm apart
on the upper right lid margin of m. orbicularis oculus, integrated by a Beckman
9852A coupler (measured time constant, 50 ms), and led through Beckman pre- and
post-amplifiers (bandpass .16-150 Hz) to an analog-to-digital converter.
To examine the possibility that ERPs were contaminated by
volume-conducted myogenic activity, recordings were made from three additional
muscles—medial frontalis, masseter and post-auricularis. Activity
from these muscles was measured with monopolar
leads, recorded via miniature electrodes (2-mm diameter Med Associates
TDE-23-48), with active electrode location for post-auricularis from an
electrode directly above the right mastoid process; for medial frontalis, from
an electrode 2 cm above the upper edge of the right supraorbital ridge and 1 cm
medial to a vertical line traversing the pupil; for the masseter muscle, from an
electrode 2 cm superior to the most lateral point on the right mandibular angle
along a line projected to the exocanthion (Fridlund & Cacioppo, 1986). EEG electrodes were located according to the International
10-20 System (Jasper, 1958) at midline frontal (Fz) and central (Cz) sites, the
right fronto-lateral site (F8), and midway between the right temporal and
temporo-parietal sites (T4/6). A
common reference electrode was placed on the right earlobe and a common ground
at FPz. Except for blink, leads
were amplified by Grass preamplifiers (bandpass .1-35 Hz) and were sampled at
1kHz for 745 ms, beginning 25 ms prior to onset of each control stimulus and the
first stimulus of each pair. Blink
was sampled at 1kHz for 250 ms beginning with onset of a reflex-eliciting tone.
Subjects were seated in a reclining chair with headrest support in an air conditioned, sound-attenuated chamber. They were instructed to remain alert, maintain eye fixation on a designated point and, to prevent jaw clenching, maintain their lips in a slightly parted position.
Each subject received four conditions: Two control conditions of low-intensity, 10-psi puff and high-intensity, 110-dB 800-Hz tone presented singly; two paired conditions of puff preceding tone by 60 and by 360 ms after adjusting for puff transit time. The four conditions occurred equally often in each of four 20-trial blocks, with order of conditions determined by a 4 x 36 matrix formed by nine 4 x 4 Latin squares. Subjects were randomly assigned without replacement to begin at a different row of the matrix. Intertrial intervals varied from 18 to 28 s (Mean = 23 s, rectangular distribution), except that a 3-minute rest period followed trial 40. To reduce the possibility of artifact on early trials, two 110-dB tones were presented before the first trial.
After each trial, subjects rated stimuli using the Stevens (1956) magnitude estimation procedure that requires numbers to be assigned to stimuli based on the estimated ratio of their loundess/intensity to anchoring standards. The standards, 90-dB tones and psychophysically matched 22-psi puffs, were delivered at the beginning of each block of trials and were assigned the rating “10.” Subjects rated each control stimulus and each pair member; subjects rated pair members in order of first then second, or second then first, with order of ratings counterbalanced over halves of the experiment. Ratings were communicated by intercom and recorded by the experimenter.
Reflex Eye Blink. A blink-scoring program (Balaban, Losito, Simons, & Graham, 1986) determined trial acceptability (3.44% discarded) and measured peak magnitude in the envelope of integrated EMG activity during 250 ms following onsets of control and S2 tones. Trials were then averaged per subject per condition. Two planned 1-df contrasts conducted on each major identified deflection tested for: 1) the presence of PPI (long SOA vs. control) and 2) refractoriness (short vs. long SOA).
ERP and EMG Waveforms. Single-trial ERPs and EMGs were subjected to eye movement correction (Gratton, Coles, & Donchin, 1983; Miller, Gratton & Yee, 1989) to adjust for effects of blink, and vertical and horizontal eye movements2.
Averages per subject, per stimulus condition, per scalp site were converted to baseline-deviated curves by subtracting the mean of the 25 prestimulus baseline points from each waveform and, because S1 activity persists beyond the S1-S2 interval puff-S1 EOG-corrected waveforms were subtracted from EOG-corrected pair waveforms (see Perlstein et al., 1993, Figure 1).
ERP components were identified and measured in the following manner: First, midline midlatency ERP components were identified using both varimax-rotated principal components analyses (PCA) derived from cross-product matrices and base-to-peak amplitude scoring methods. For the PCA, inputs were the first 85-ms samples from the waveforms for 36 subjects, 2 leads (Fz, Cz) and 3 SOA conditions (tone-alone and S2 tones at 60 ms and 360 ms). Because they are not associated with actual ‘peaks’, points chosen at the window extremes (Table 1) were excluded3.
Later-latency components were scored using base-to-peak amplitudes measured within windows (Table 1) for all midline (N100, P200) and lateral components (C1, C2)4.
EMG data were also measured by measuring base-to-peak amplitudes within specified windows, but without exclusion of points chosen at window extremes.
Pairing effects on all ERP components and EMG deflections were evaluated using two 1-df planned contrasts to determine if: 1) the long-SOA response exhibited significant PPI (long SOA vs. control) and 2) if inhibition was greater at the long than short SOA (refractoriness). In addition, the three N100 components were evaluated by scaling, per subject/site/SOA, using the range-correction method ([xi-minimum] / [maximum – minimum]). These scaled data were then evaluated by 3-Component x 3-Lead ANOVAs and for differences in linear and quadratic change over the three SOAs. The midline P30 and P50 were similarly range corrected and analyzed using a 2-Component x 3-SOA ANOVA and for differences in linear and quadratic change over the three SOAs.
Magnitude Estimates. Per subject averages were computed for the 10 ratings made under each of the 12 conditions—2 instructions (pair members in order first then second, or second then first) x 2 stimuli x 3 SOAs. Collapsed across instructions, the ratings of S1 (backward effects of S2) and S2 (forward effects of S1) were analyzed separately by 3-SOA ANOVAs and follow-up contrasts. The Greenhouse and Geisser (1959) epsilon adjustment was applied to correct for the use of repeated-measures with more than two levels of a factor. Post-hoc contrasts used the Studentized Range (Q) statistic to control for multiple comparisons.
![]()
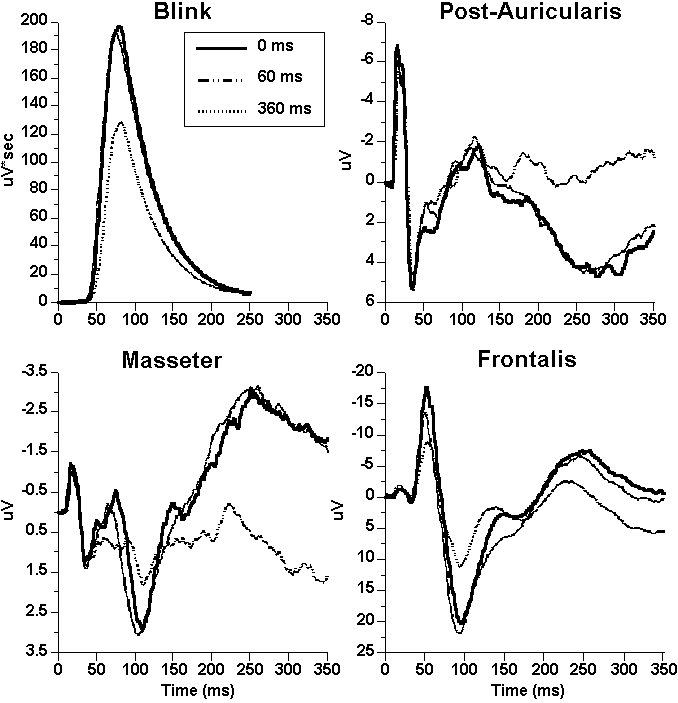
Reflex blink. Effects of puff-tone pairing on blink are shown in the waveforms of Figure 1 and mean magnitude provided in Table 2. Table 3 provides the planned statistical contrasts. Puff-tone pairing significantly reduce blink magnitude at the long SOA compared to both the short SOA and control conditions.
Masseter, post-auricularis, medial frontalis. Waveforms for EOG-corrected scalp and facial muscles are also shown in Figure 15.
Consistent with the literature, each comprised initial biphasic negative
(N)-positive (P) responses within the first 30 ms and neither masseter nor PAR
were significantly affected by pairing for either deflection.
On the other hand, medial
frontalis showed several significant modulations in the midlatency region: N19
was enhanced over the control at the long SOA; N40 was smaller at the long SOA
than both the control and short SOA (Tables 2 and 3).
Later responses for all three
muscles were generally reduced at the long compared to the short SOA, though not
significantly in all cases.
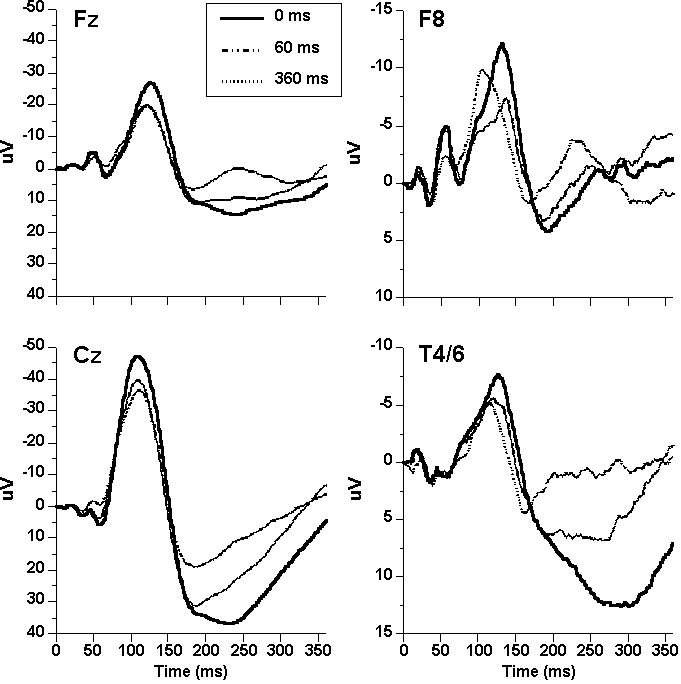
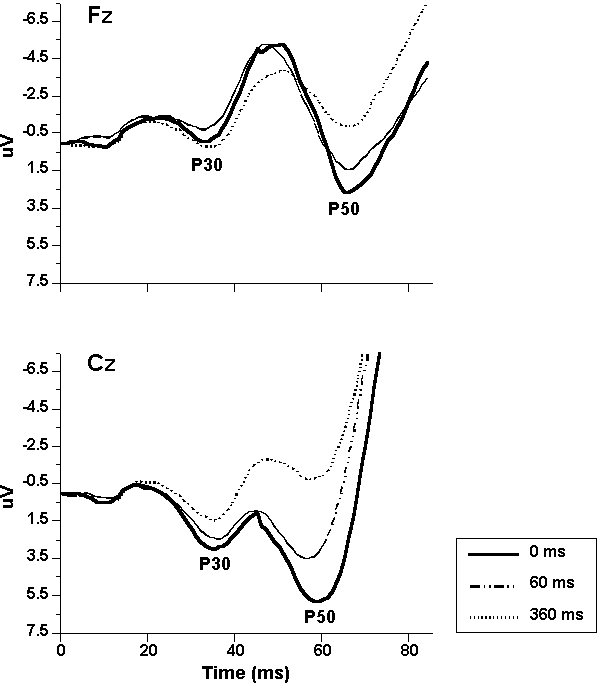
Midlatency ERPs. Grand mean EOG-corrected midline ERPs (Figure 2, left) contained the following mid- and later-latency deflections: N19, P30, N40, P50, N100, and P200. N19 and N40 were largest at Fz; all other midline components were largest at Cz. The amplitudes of the ERPs elicited by acoustic S2s, beginning with N40, were reduced by tactile S1s and, like effects on blink, reductions were generally greater at the longer SOA. In contrast, N19 and P30 were not significantly affected by pairing at either SOA. Pairing effects on midline, midlatency ERPs are shown more clearly in Figure 3, illustrating the first 85-ms waveforms at midline.
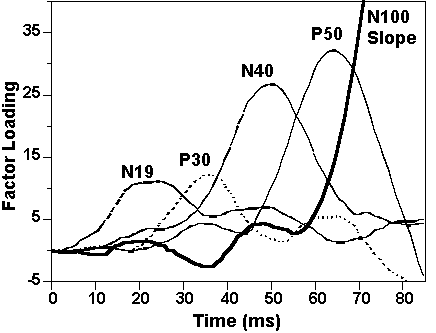
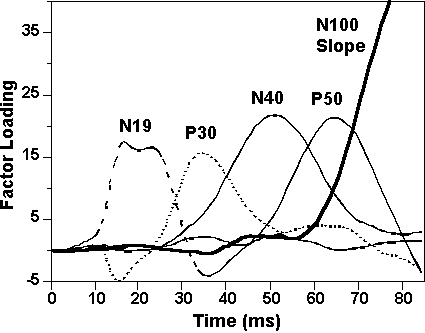
The first five of 10 factors from the PCA on the 85-ms epoch of the midline leads (Figure 4) accounted for 98.7% of the total variance in the data set. The maximum loadings of these factors suggests identification of factors 4, 5, 2, 3, and 1 with N19, P30, N40, P50, and the rising slope of N100, respectively. Mean factor scores are shown in Table 4. Planned contrasts (Table 5) confirmed the illustrated pairing effects on P50 showing that, at both midline sites, it was significantly reduced below control levels at the long SOA and at the long compared to short SOA. Thus, contrary to the refractory hypothesis, P50 inhibition was significantly greater at the long than short SOA at both leads. Factor scores associated with the earlier N19 and P30 were again not significantly affected by pairing at either SOA for either contrast. This pattern of statistical effects persisted after correction of ERPs for myogenic activity6.
Effects of pairing may have begun as early as N40, which was reduced at Fz and enhanced at Cz at the long SOA compared to the control condition. N40 was contaminated by volume-conducted frontalis muscle activity, however, and therefore will not receive further consideration. Means for the supplementary peak amplitude measure are provided in Table 4. Analyses (Table 5) replicated the PCA results with respect to the most critical questions; namely, the absence of significant pairing effects on P30, and greater inhibition of P50 at the long SOA compared to control and at long compared to the short SOA.
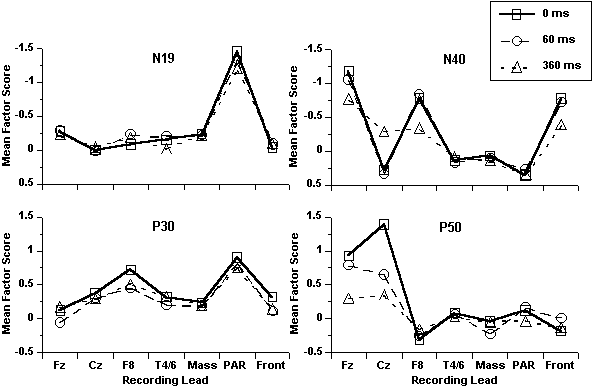
Myogenic activity and midlatency ERPs. To explore the possibility that the midlatency ERP findings described above were appreciably and systematically influenced by myogenic activity, a set of analyses was conducted based on the topographical distribution of deflections in the midlatency range. Specifically, a PCA was performed on a data set comprising the first 85 ms of EOG-corrected data from all ERP and muscle recording sites. The first five of 10 factors extracted accounted for 97.0% of total variance in the data set and ordering of the maximum loadings suggests identification with N19, P30, N40, P50, and the rising slope of N100 (Figure 5). Plots of the mean factor scores as a function of lead and SOA (Figure 6) show that N19 and P50 were strictly localized to particular sites: N19 was maximal over PAR and only slightly negative at all other leads; P50 was restricted to the two midline sites (Fz, Cz). The other two components, P30 and N40, had peaks at both muscle and ERP sites. However, because response at the electrode closest to the mastoid (i.e., T4/6) was not elevated, it is unlikely that P30 measured at the midline was contaminated by volume conduction from the PAR. In contrast, N40 peaked at all three frontal sites—Fz, F8, and frontalis—suggesting that it was particularly vulnerable to volume-conducted frontalis activity.
Analyses of pairing effects on the midlatency factor scores derived from this 7-lead PCA (Table 6) showed that there were no significant SOA effects on N19 and P30 at any ERP recording site, while P50 was significantly inhibited, and more so at the long that short SOA. In contrast, two significant frontalis effects not present in the EMG data described earlier (Figure 4)—reductions in P30 at both SOAs and facilitations of P50 at the short SOA—differ from effects on the corresponding ERPs described below, strongly suggesting that these midline ERP components were unaffected by volume-conducted frontalis activity.
Later-latency ERPs. Pairing effects on the midline late exogenous ERPs (N100, P200; Figure 2, Table 4) closely paralleled the effects on P50. Planned comparisons (Table 5) confirm that prestimulation significantly inhibited N100 at the long SOA compared to control at both midline sites, and at the long compared to the short SOA at Cz. Planned comparisons on P200 showed that reductions were significant at both leads for the control vs. long SOA and short vs. long SOA comparisons. These effects were unchanged by EMG corrections except that, after correction for frontalis, the N100 reduction at Fz also became significantly greater at the long than short SOA.
Visible in the averaged waveforms at F8 and T4/6 (Figure 2) are the two specific N100 components—C1, most prominent at F8, and C2 (Tb), present at both F8 and T4/6. An early deflection between Component s 1 and 2—Ta of the T-complex—can also be seen in some conditions at both leads. Although these components are generally visible only as inflection points in the grand mean ERPs, they are clearly visible in the majority of individual-subject averages. Mean base-to-peak amplitudes for these components are shown in Table 4. Planned contrasts (Table 5) on C1 revealed it was significantly reduced only at F8 in the short vs. long SOA comparison. In contrast, C2 was significantly reduced at the long-SOA compared to control at F8 and, at T4/6, at the long SOA vs. control at T4/6.
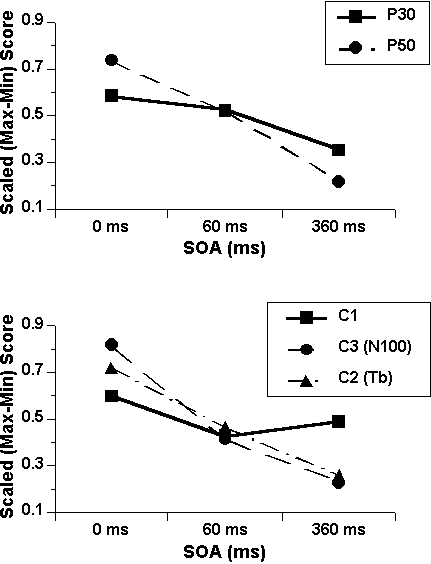
As described above, differential sensitivity to pairing was noted between the two midline positive ERP components, P30 and P50, and among the three negativities that comprise the N100. The pattern of these effects is consistent with the hypothesis that nonspecific auditory ERP components show prepulse inhibition, while the specific ERP components are much less, if at all, affected by the presence of a prepulse. However, a direct comparison among components requires equating for their large differences in size: P50 is much larger than P30 and the nonspecific (Component 3) N100 is much larger than either of the two specific components. Thus, to make this direct comparison and to make it with maximum sensitivity, baseline-to-peak amplitudes were range corrected according to the formula [(xi - max) / (max - min)] applied to the set of SOAs per subject. The midlatency-midline components and the three N100 components were analyzed separately. P30, P50, and the nonspecific N100 were scaled at the Cz electrode site where they were maximal and most sensitive; the specific N100s (C1 and C2) were scaled at T4/6 where their overlap with C3 was minimal (Loveless & Brunia, 1990) and where they were farthest from possible influence by the medial frontalis P90 response. Mean scaled data are presented in Figure 5. ANOVAs on the two data sets evaluated differences among components in linear and quadratic changes over SOA. Consistent with the top panel of Figure 7, analysis of the two midlatency components—P30 and P50—revealed a significant linear trend over SOA (F[1,35] = 13.11, p <. 001) and, more importantly, a significant linear SOA X Component interaction (F[1,35] = 10.57, p < .003). Follow-up trend tests on each component separately, conducted to maximize power at the cost of an increased alpha, revealed a significant linear effect of SOA on the nonspecific, P50, component (F[1,35] = 25.81, p < .0001); P30 remained relatively insensitive to the delivery of the prepulse at either SOA.
Figure
7 (bottom panel) likewise suggests that the three N100 components differed
in their linear change from control levels to levels at the 360-ms SOA.
This was confirmed statistically by a significant linear SOA X Component
interaction (F[1,35] = 8.44, p < .0015).
More importantly, the two specific components differed from the
nonspecific component, as reflected in a significant quadratic effect of
component (C1 vs, C2 and C3) on the linear trend over SOA (0- vs. 360-ms; F[1,
35] = 7.26, p <. 01),
primarily due to the faster recovery of C1 compared to C2 and C3. Post-hoc linear SOA X Component trend tests with Bonferoni-adjusted
p-values (.05/2 = .025) showed that C1 differed from both C3 (N100) and
C2 (Fs[1, 35] ≥ 10.06, ps < .003).
As in Perlstein et al. (1993), pairing significantly affected judgments of both pair members compared to the same stimuli presented alone. Mean tone ratings were 21.25, 19.90 and 18.13 for the 0-, 60- and 360-ms SOA conditions, respectively. Mean puff ratings were 6.96, 8.24 and 7.75 for the 0-, 60- and 360-ms SOA conditions, respectively. Relative to control-tone judgments, puff significantly reduced the loudness of S2 tone ratings compared to ratings of the tone presented alone. Tone judgments were significantly reduced at both the short and long SOA compared to the control (Qs[35] = 7.43 and 15.31, respectively, ps < .01), and paired condition ratings significantly differed from one another (Q[35] = 9.86, p < .01). Backward effects of tone on S1-puff judgments were also significant (F[2, 70] = 18.76, p < .001); pairing enhanced judgments of puff intensity at the short and long SOAs (Qs[35] = 6.78 and 7.27, respectively, ps < .01), and the two paired conditions differed from one another (Q[35] = 3.54, p < .05).
![]()
The primary goals of the present research were to replicate and extend the findings of Perlstein et al. (1993) and evaluate the hypotheses that: 1) prestimulus-induced midlatency ERP reductions are, like blink PPI, due to an extrinsic mechanism; 2) blink PPI is associated with differential effects on modality specific and nonspecific ERP components; and 3) PPI is associated with perceptual concomitants consistent with suggestions that it reflects an underlying sensory protective mechanism. An additional, exploratory goal was to assess effects of prestimulation on later-latency specific and nonspecific ERP components.
Two key findings were obtained concerning crossmodal prestimulation effects on ERPs. First, the amplitudes of P50 and later midline components elicited by a startle-intensity acoustic S2 were reduced when shortly-preceded by a low-intensity S1. These modulatory effects are opposite those that would be predicted if reductions are due solely to an intrinsic or passive refractory-type mechanism. Second, P30 and P50 were differentially affected by prestimulation, replicating the findings of Perlstein et al. (1993) for acoustic pairs.
The present findings also demonstrated that later-latency specific and nonspecific components of the N100 were differentially affected by prestimulation. The midline N100, which predominantly reflects Component 3 and activity within the modality nonspecific system, and the Tb wave of Component 2, which presumably reflects late activity within the modality specific system, showed prestimulus effects that essentially paralleled those of P50 as a function of SOA. That is, both were reduced by tactile prestimulation, and reductions were greater at the longer SOA. In contrast, the earlier Component 1 showed a tendency toward inhibition at the short SOA and recovery towards baseline at the longer SOA. This finding functionally dissociates the nonspecific and the earlier of the specific N100 components.
The present experiment also replicated the perceptual findings, under different stimulus conditions, of Perlstein et al. (1993). The perceptual magnitude of a low-intensity S1 was enhanced while that of a high-intensity S2 was reduced relative to control stimuli: S1 enhancement was greater at the short than long SOA; S2 reduction was greater at the long than short SOA. These data are consistent with the hypothesis that the mechanism underlying blink PPI and prestimulus reductions of nonspecific ERPs indexes a preattentive protective mechanism that facilitates the processing to recognition of a low-intensity S1 in the face of competing or potentially disruptive impact of a high-intensity S2.
Below
we consider the nature of the PPI-like effects on mid- and later-latency ERP
deflections, potential myogenic influences on ERP findings, and pairing effects
on judgements of stimulus intensity.
Nature of Prestimulus Inhibitory Effects on Midline ERPs.
The findings that midline P50, N100, and P200 elicited by acoustic S2s
are reduced by tactile S1s and that this PPI effect is delayed (i.e., reduction
is greater at the longer SOA), suggest that the inhibitory effect
involves an active extrinsic mechanism; that is, modulation by a path external
to but synapsing with the path that itself generates the potential.
This suggestion contrasts with previous assertions that ERP decrements
result from a passive intrinsic mechanism, such as short-term exhaustion of
excitatory processes inherent in the neural aggregates which mediate the
response (e.g., Callaway, 1973; Naber, Kathman & Engel, 1992; Roemer,
Shagass, & Teyler, 1984; Webster & Aitkin, 1971).
An intrinsic or refractory-type mechanism is typically observed in
studies of stimulus rate effects, wherein response amplitude decreases with
increased stimulation rate (e.g., Bess & Ruhm, 1972; Butler, 1973; Davis,
1968; Davis, Mast, Yoshie & Zerlin, 1966; Nelson & Lassman, 1968; Roth
et al., 1976; Rothman, Davis, & Hay, 1970; cf., Budd & Michie, 1994). The
present study's use of a long (18 to 28 s) interpair interval eliminated
contamination by rate effects, at least for the midlatency ERPs and for
Component 1, given that it is well within the recovery time required for these
components. The nonspecific
component, in contrast, may require as long as 2 minutes for full recovery
(e.g., Fruhstorfer, 1971; Näätänen & Picton, 1987) and recovery time for
Tb is unknown.
Two parameters—pair-member modality and SOA—are central in any attempt to differentiate between refractory and active inhibitory processes in ERP components. As mentioned in the introduction, the use of ipsimodal-pair members confounds refractoriness due to modality-specific effects at peripheral levels of the input path with other forms of modulation, such as PPI (Balaban et al., 1985; Rimpel et al., 1982). Davis et al. (1972) found that reductions in N100 and P200 were, on average, 50% greater with ipsimodal than with crossmodal pairs, supporting this suggestion. Regarding SOA, intrinsic and extrinsic processes can not be dissociated, regardless of pair-member modality, when the shortest SOA exceeds that which produces maximal inhibition. Under this condition, the response amplitude-SOA function would reflect a monotonic increase associated with the recovery limb because refractory effects are greatest at near-simultaneity of two stimuli and monotonically decrease with increasing SOAs. This is precisely what was reported by Nagamoto, Adler, Waldo and Freedman (1989) who employed paired acoustic stimuli delivered at intervals of 75, 150, and 500 ms and obtained a monotonic increase in P50 amplitude as a function of increased SOA. Again, when pair-member modality and SOA are unconfounded as they were in the present study, effects of extrinsic and intrinsic processes can be more clearly ascertained.
Differential Prestimulus Effects on Midline Midlatency ERPs. The present findings suggest that pairing differentially affects P30 and later midline potentials, with P30 not showing a statistically significant pairing effect and later midline components showing significant amplitude reductions at both SOAs. Perusal of Figure 3, however, suggests that there may have been some inhibition of P30 at the long SOA and raises the possibility that our procedures were not sufficiently sensitive to detect it. Although a more definitive answer must await further research, there are several factors that mitigate against a lack-of-sensitivity argument. First, pairing effects on P30 were in opposite directions at the Fz and Cz recording sites; this is consistent with our statistical conclusion of random variation. Second, the differential pairing effects on P30 and P50 replicate precisely those observed in our previous study (Perlstein et al., 1993) and those reported by Adler, Pachtman et al. (1982). Third, the P30 and P50 components were quantified both as base-to-peak measures and through factor analytic techniques. The differential pairing effects were consistent across the two methods. This was true in Perlstein et al. (1993) as well. Finally, the differential effects of pairing on P30 and P50 are consistent with the alignment of these two midlatency components with later-latency N100 components associated with modality specific and nonspecific projection systems which themselves were not (Component 1) and were (Component 3) sensitive to pairing.
Modality specific and nonspecific information is transmitted in parallel, asynchronously, from midbrain to higher levels via rapid- and slow-conducting systems, respectively. Activity arising from auditory stimuli is transmitted by the rapidly-conducting specific, or classical lemniscal path. This pathway originates in the cochlea and extends through the central nuclei of inferior colliculi to the ventral division of the medial geniculate and then to primary auditory cortex where N19 and P30 are generated (Hashimoto et al., 1995; Liégeois-Chauvel, Musolino, & Chauvel, 1991; Liégeois-Chauvel, Musolino, Badier, Marquis, & Chauvel, 1994; Scherg, Hari, & Hämäläinen, 1989). Activity arising from auditory and tactile stimuli is also transmitted by the more slowly-conducting nonspecific (extralemniscal) path generating P50 and later nonspecific potentials (Erwin & Buchwald, 1986a). This pathway corresponds to the medial projection system described by Erwin and Buchwald (1986a), or the “lemniscal adjunct” system described by Weinberger and Diamond (1987, 1988; see also Di & Barth, 1993) and comprises a parallel conduction route centered on the medial and dorsal divisions of the medial geniculate. This nonspecific auditory system ascends medially through the reticular formation, including polysensory cuneiform nuclei, and to midbrain structures that do not receive direct ascending lemniscal-line input. It branches from the lemniscal path to the polysensory extralemniscal path at the level of the inferior colliculus (Aitkin, 1986; Weinberger & Diamond, 1987), which may be the earliest level at which cross-modal interactions occur in the auditory system (Szczepaniak and Møller, 1993), and thence projects broadly to secondary auditory cortex and association areas. It is generally believed that prestimulus-intitiated activity feeds into the direct startle path at the level of the cuneiformis reticularis nuclei and inhibits the startle reflex for periods from about 15 to 400 ms or more (e.g., Graham, 1975), by efferent modulatory activity from several structures, such as portions of the tegmental nuclei (e.g., Koch & Schnitzler, 1997) and ventral pallidum (Kodsi & Swerdlow, 1997).
Thus, existing functional and neuroanatomical data suggest that P30 and P50 do not depend on serial transmission via the specific projection system but involve parallel asynchronous paths which may transmit information about transient and sustained stimulus characteristics, respectively. Additionally, as Perlstein et al. (1993) suggested, the cortical areas involved in early sensory-perceptual processing appear to receive in parallel at least two kinds of stimulus information: Information that is independent of a preceding stimulus and information that is not. This suggestion is consistent with the position that the specific sensory nuclei of the thalamus transmit “discriminative” aspects of ascending sensory information that are relayed to the cerebral cortex, while information relayed by midline-intralaminar nuclei to more diffuse areas of cortex and basal ganglia contains “nondiscriminative” information (Groenewegen & Berendse, 1994). The present findings also provide evidence that P30 is generated in the specific path at a point prior to sensory convergence; P50 is generated at a point beyond which somatosensory and acoustic stimulus-processing converge.
Specific versus Nonspecific N100 Components. Although the Perlstein et al. (1993) hypothesis concerned only a specific and nonspecific system, the present findings, together with the literature, may support three rather than two systems. That is, Components 1 and 3 reflect activity from the specific and nonspecific systems, respectively; Tb of Component 2 exhibits functional characteristics of an intermediate system, perhaps involving the "lemniscal adjunct" pathway described by Weinberger and Diamond (1987).
The differential pairing effects on Components 1 and 3—rapid recovery toward baseline for Component 1 and increased inhibition of Component 3 at the long SOA—supports previous work suggesting both their functional and anatomical independence. Other functional dissociations between these components shown by other studies include differences in the effects of stimulus intensity and stimulation rate: The amplitude of the magnetically-recorded N100 (N100m; Component 1) reached a plateau at 60 dB, while the electric N100, recorded at one-half the distance up from T4 to C4 and presumably reflecting Component 2 or 3, or both, continued to increase linearly up to 100 dB (Reite, Zimmerman, Edrich, & Zimmerman, 1982). Other studies that have recorded ERPs at Cz (Picton, Goodman, & Bryce, 1970; Roth, Dorato, & Kopell, 1984) have also observed increases in N100 amplitude up to approximately 110 dB. The Component 1, reflected in N100m, has also been shown to be less sensitive to stimulation rate than the vertex-recorded electrical N100 (Component 3), consistent with rapid recovery of specific-system neurons (Hari, Kaila, Katila and Varpula, 1982); that is, the vertex-recorded electrical N100 reached a plateau at an ISI of 4 s, while Component 3 amplitude continued to increase up to the 16-s the maximum ISI. Furthermore, there is evidence, consistent with the present study, that N100m is unaffected by crossmodal paired stimulation (Huttunen, Hari and Vanni, 1987), and may even be enhanced by ipsimodal acoustic pairs (Loveless, Hari, Hämäläinen, & Tiihonen, 1989; see also Loveless & Hari, 1993).
Prestimulus reductions of Component 2 in the present study, which paralleled reductions of Component 3, indicate that the pathway generating this response likely receives input from a somatosensory-processing source. Findings of attention-related enhancements of Component 2 (i.e., Tb; Dien, Tucker, Potts & Hartry-Speiser, 1997; Hackley, Woldorff & Hillyard, 1990; Perrault & Picton, 1984), presumably mediated by the nonspecific system, because attention effects are not restricted to a single modality, also suggest overlapping functional aspects of Components 2 and 3. However, Loveless and Brunia's (1990) finding that effects of increased stimulus rise-time (increased latency and decreased amplitude) were restricted to the nonspecific Component 3 indicates that Component 2 also exhibits functional characteristics of the specific system.
Thus, anatomical and behavioral evidence are in agreement for Component 3 and for Component 1 but findings for Component 2 suggest that this component exhibits functional and anatomical characteristics of an intermediate system. The question of whether or not the specific N100 components are contaminated by volume-conduction from Component 3 also needs to be considered. This possibility was noted by Loveless and Brunia (1990) who, consequently, measured both components 1 and Tb at a temporal site, despite the F8 maximum for Component 1. Perrault and Picton (1984) noted that both Components 1 and 2 were maximal at temporal sites. Given the decreased likelihood of Component 1 and 2 overlap with Component 3 at a temporal site, as well as functional differences in the three components, it is unlikely that any volume-conduction from Component 3 could account for the observed pairing effects at this lead.
EMG/ERP relationships. The high-intensity stimuli employed in this research elicited myogenic responses that were evident earlier than 50 ms, and during the latency range of the N100 components. However, several points suggest that myogenic influence, although present, did not underlie either the inhibitory effects of prestimulation on midline P50 and later potentials or the absence of pairing effects on P30. First, we analyzed ERPs following removal of EMG contamination by correcting ERPs for each individual muscles singly and for all three muscles together using the Gratton et al. (1983; Miller et al., 1989) algorithm6. Following this procedure, P30 remained statistically unaffected by pairing and P50 inhibition remained significant. The pattern of statistical effects for the later potentials was also unchanged. Second, the spatial distribution of factor scores obtained from the PCA conducted on midlatency responses in the four EEG and three EMG leads (Figure 6), at least for the midlatency ERP deflections of primary interest—midline P30 and P50—is not spread across the scalp in a manner consistent with volume conduction to the midline. Finally, the effects of pairing on early muscle responses, although nonsignificant, were not even in the same direction as the effects on ERPs.
In
contrast to the midline P30 and P50, myogenic influence on N40 was more
pronounced. The PCA on combined
midlatency ERP and EMG potentials extracted a large, temporally-extended factor
loading on the N40 region (Figure 5) whose factor scores were large for the
frontalis muscle and for the two frontal ERP leads, Fz and F8 (Figure 6). This myogenic contamination was not evident at the more
posterior recording sites (Cz and T4/6). Frontalis
also has a large response at P90 and might, therefore, be expected to
contaminate the later-latency N100 components at the two frontal leads.
Examination of the components before and after correction of ERPs for
frontalis muscle activity suggests that contamination did occur, as evidenced by
increases in negative potentials after correction of frontal leads.
However, at T4/6, the amplitude changes were again near zero; at Cz, the
changes ranged from 1.4 to 3.5 mV compared to 1.7 to 5.7 mV at Fz.
Further, the significance pattern of SOA effects was the same for
corrected frontal and non-frontal leads, suggesting that the correction was
effective.
The findings of enhanced magnitude estimates of S1s and reduced estimates of S2s replicate the intramodal pairing effects of Perlstein et al. (1993) and Filion and Ciranni (1994) and, for reductions of S2, also the crossmodal findings of Cohen, Hoffman and Stitt (1981). Furthermore, these findings are consistent with the hypothesized protective function of the mechanism underlying prestimulus modulatory effects on blink and ERPs. However, as noted by Perlstein et al. (1993), and not mutually exclusive of the “protection” hypothesis, the perceptual modifications obtained may be seen as assimilation or context effects: Perceptual magnitudes of the pair members shifted toward one another, exhibiting a central tendency effect. Such context effects are thought to occur at the perceptual rather than the judgmental stage of processing (e.g., Algom & Marks, 1990; Marks, 1988; Marks, Szczesuil & Ohlott, 1986; Marks & Warner, 1991; Parker & Schneider, 1994; Rankin & Marks, 1991; Schneider & Parker, 1990) and, therefore, are due, in part, to modification of the underlying perceptual representation of stimuli, rather than the overt response per se.
It is clear, however, that the relationship between perceived stimulus intensity and PPI is complex. Whether a protective function is conferred by or observed to be associated with prepulse inhibition appears to be dependent upon several critical factors. First, when stimulus pairs are of unequal intensity assimilation effects might be inseparable from other forms of perceptual modulation. The findings from the present study and from Perlstein et al. (1993) demonstrate the robustness of these assimilation effects (see also, Blumenthal, Schicatano, Chapman, Norris & Ergenzinger, 1996; Filion & Ceranni, 1994). Second, the strength of these assimilation effects raises the question of whether judgements of perceptual magnitude or intensity provide the most sensitive method for assessing PPI-concomitant perceptual influences (e.g., Norris & Blumenthal, 1996). Third, there is a question about whether or not the motor response associated with startle can be entirely dissociated from judgements of perceptual magnitude; that is, whether “the subjective perception of the startle stimulus is strongly influenced by the magnitude of the startle reflex elicited by that stimulus” (Swerdlow, Geyer, Blumenthal & Hartman, 1999, p. 544). Associations between the strength of blink PPI and perceived loudness of the startle-eliciting stimulus have been demonstrated (e.g., Filion & Ceranni, 1994), though clear dissociations exist (e.g., Swerdlow et al., 1999). One experimental approach that has bolstered support for the protection-of-processing hypothesis is relating the accuracy of detection of a prepulse to the magnitude of startle inhibition. Norris and Blumenthal (1996) used such a method and did demonstrate enhanced prepulse detection accuracy on trials that showed PPI compared to trials that did not.
Finally, although the absence of reduction in S1 is compatible with an hypothesized mechanism protecting its processing from disruption (Graham, 1975, 1979; Perlstein et al., 1993), pairing effects on blink, ERPs and perception of S2 were dissociated in the Perlstein et al. studies. That is, in a condition where stimulus-pair members were of low intensity (75 dB), S2 judgments showed an increase, while ERPs to S2 decreased. However, such dissociations are not inexplicable and do not necessarily contradict the sensory-protective hypothesis. As noted by Perlstein et al. (1993), exogenous ERPs, blink, and PPI reflect primarily the transient aspects of stimulation (Berg, 1973; Graham, 1975; Onishi & Davis, 1968). In contrast, perception of stimulus intensity depends mainly on the partial summation of steady-state portions of the stimulus envelope (Gerken, Bhat, & Hutchison-Clutter, 1990). Thus, perception of intensity presumably reflects sustained neural activity that is capable of maintaining long-lasting representations of a stimulus. The ERPs obtained in the present research are largely insensitive to steady-state aspects of stimulation, unlike the sustained potential elicited by stimuli of longer duration (e.g., Hari, Aittoniemi, Järvinen, Katila, & Varpula, 1980; Picton, Woods, & Proulx, 1978a, 1978b) which more faithfully reflects both stimulus intensity and duration (i.e., total stimulus energy).
The
principal findings of the present research are the demonstrations that 1) P50
inhibition is not due to a passive refractory mechanism, but instead to an
active inhibitory process, and 2) P30 and P50 are differentially affected by
prestimulation. These findings have
particular relevance to the study of schizophrenia (e.g., Adler et al., 1982)
and other disorders in which “sensory gating” abnormalities have been
hypothesized (e.g., post-traumatic stress disorder; Neylan et al., 1999), and in
which reduced P50 inhibition to paired acoustic stimuli has been observed.
Absent from these studies have been attempts to differentiate between
intrinsic and extrinsic mechanisms, as only ipsimodal stimulus pairs have been
employed. Several studies of
schizophrenia have used multiple interstimulus intervals (Nagamoto et al., 1989;
Nagamoto, Adler, Waldo, Griffith & Freedman, 1991), and showed monotonic
increases in P50 amplitude with increased intervals.
However, the present findings encourage the use of cross-modal pairs and
a selected range of pairing intervals in future studies of psychopathology in
order to deconfound intrinsic and extrinsic inhibitory processes and more fully
understand the neural mechanisms of PPI dysfunction.
Adler, L. E., Pachtman, E., Franks, R. D., Pecevich,
M., Waldo, M. C., & Freedman, R. (1982). Neurophysiological evidence for a
defect in neuronal mechanisms involved in sensory gating in schizophrenia. Biological
Psychiatry, 17, 639-654.
Aitkin, L. (1986). The Auditory Midbrain: Structure
and Function in the Central Auditory Pathways. New Jersey: Humana Press.
Algom, D., & Marks, L. E. (1990). Range and
regression, loudness scales, and loudness processing: Toward a context-bound
psychophysics. Journal of Experimental Psychology: Human Perception and
Performance, 16, 706-727.
Balaban, M. T., Anthony, B. J., & Graham, F. K.
(1985). Modality-repetition and attentional effects on reflex blinking in
infants and adults. Infant Behavior and Development, 8, 443-457.
Balaban, M. T., Losito, B., Simons, R. F., &
Graham, F. K. (1986). Off-line latency and amplitude scoring of the human reflex
eye blink with Fortran IV. Psychophysiology, 23, 612 (Abstract).
Berg, K. M. (1973). Elicitation of acoustic startle in
the human. Unpublished doctoral thesis, University of Wisconsin, Madison.
Bess, J. C., & Ruhm, H. B. (1972). Recovery cycle
of the acoustically evoked potential. Journal of Speech and Hearing Research,
15, 507-517.
Blumenthal, T. D., & Gescheider, G. A. (1987).
Modification of the acoustic startle reflex by a tactile prepulse: The effects
of stimulus onset asynchrony and prepulse intensity. Psychophysiology, 24,
320-327.
Blumenthal, T. D., & Tolomeo, E. A. (1989).
Bidirectional influences of vibrotactile stimuli on modification of the human
acoustic startle reflex. Psychobiology, 17, 315-322.
Blumenthal, T. D., Schicatano, E. J., Chapman, J. G.,
Norris, C. M., & Ergenzinger, E. R. (1996). Prepulse effects on magnitude
estimation of startle-eliciting stimuli and startle responses. Perception
& Psychophysics, 58, 73-80.
Budd, T. W., & Michie, P. T. (1994). Facilitation
of the N1 peak of the auditory ERP at short stimulus intervals. Neuroreport,
20, 2513-2516.
Butler, R. A. (1973). The cumulative effects of
different stimulus repetition rates on the auditory evoked response in man. Electroencephalography
and Clinical Neurophysiology, 35, 337-345.
Callaway,
E. (1973). Habituation of averaged evoked potentials in man. In H. V. S. Peek
& M. J. Herz (Eds.), Habituation: Volume II. Physiological substrates
(pp. 153-174). New York: Macmillan.
Celesia, G. G. (1976). Organization of auditory
cortical areas in man. Brain, 99, 403-414.
Chatrian, G. E., Goff, W. R., Picton, T. W., Coats, A.
C., Celesia, G. G., & Cracco, J. B. (1984). American electroencephalographic
society guidelines for clinical evoked potential studies. Journal of Clinical
Neurophysiology, 1, 3-53.
Cohen, M. E., Hoffman, H. S., & Stitt, C. L.
(1981). Sensory magnitude estimation in the context of reflex modification. Journal
of Experimental Psychology: Human Perception and Performance, 7,
1363-1370.
Davis, H. (1968). Auditory responses evoked in the
human cortex. In A. V. S. de Reuck, & J. Knight (Eds.), Ciba Foundation
Symposium on Hearing Mechanisms in Vertebrates (pp. 259-268). London: J.
& A. Churchill.
Davis, H., Mast, T., Yoshie, N., & Zerlin, S.
(1966). The slow response of the human cortex to auditory stimuli: Recovery
process. Electroencephalography and Clinical Neurophysiology, 21,
105-113.
Davis, H., Osterhammel, P. A., Wier, C. C., &
Gjerdingen, D. B. (1972). Slow vertex potentials: Interactions among auditory,
tactile, electric, and visual stimuli. Electroencephalography and Clinical
Neurophysiology, 33, 537-545.
Davis, M. (1984). The mammalian startle response. In
R. C. Eaton (Ed.), Neural mechanisms of startle behavior (pp. 287-351).
New York: Plenum.
Deiber, M. P., Ibañez, V., Bastuji, H., Fischer, C.,
& Mauguiére, F. (1989). Changes of middle latency auditory evoked
potentials during natural sleep in humans. Neurology, 39, 806-813.
Di, S., & Barth, D. S. (1993). Binaural vs.
monaural auditory evoked potentials in rat neocortex. Brain Research, 630,
303-314.
Dickerson, L. W. & Buchwald, J. S. (1991).
Midlatency auditory-evoked responses: Effect
of scopolamine in the cat and implications for brain stem cholinergic
mechanisms. Experimental Neurology, 112, 229-239.
Dien, J., Tucker, D. M., Potts, G., &
Hartry-Speiser, A. (1997). Localization of auditory evoked potentials related to
selective intermodal attention. Journal of Cognitive Neuroscience, 9,
799-823.
Erwin, R., & Buchwald, J. S. (1986a). Midlatency
auditory evoked responses: Differential recovery cycle characteristics. Electroencephalography
and Clinical Neurophysiology, 64, 417-423.
Erwin, R., & Buchwald, J. S. (1986b). Midlatency
auditory evoked responses: Differential effects of sleep in the human. Electroencephalography
and Clinical Neurophysiology, 65, 383-392.
Erwin, R., & Buchwald, J. S. (1987). Midlatency
auditory evoked responses in the human and the cat model. In R. Johnson, J. W.
Rohrbaugh, & R. Parasuraman (Eds.), Current trends in event-related
potential research (pp. 461-467). Amsterdam: Elsevier.
Fendt, M., & Koch, M. (1999). Cholinergic
modulation of the acoustic startle response in the caudal pontine reticular
nucleus of the rat. European Journal of Pharmacology, 370,
101-107.
Filion, D. L., & Ciranni, M. (1994). The
functional significance of prepulse inhibition: A test of the protection of
processing theory. Psychophysiology, 31, S46 (supplement).
Filion, D. L., Dawson, M. E., & Schell, A. M.
(1993). Modification of the acoustic startle-reflex eyeblink: A tool for
investigating early and late attentional processes. Biological Psychology,
35, 185-200
Filion, D. L., Dawson, M. E., & Schell, A. M.
(1998). The psychological significance of human startle eyeblink modification: A
review. Biological Psychology, 47, 1-43.
Fridlund, A. J., & Cacioppo, J. T. (1986).
Guidelines for human electromyographic research. Psychophysiology, 23,
567-589.
Fruhstorfer, H. (1971). Habituation and dishabituation
of the human vertex response. Electroencephalography and Clinical
Neurophysiology, 30, 306-312.
Gerken, G. M., Bhat, V. K. H., &
Hutchison-Clutter, M. (1990). Auditory temporal integration and the power
function model. Journal of the Acoustical Society of America, 88,
767-778.
Graham, F. K. (1975). The more of less startling
effects of weak prestimulation. Psychophysiology, 12, 238-248.
Graham, F. K. (1979). Distinguishing among orienting,
defense, and startle reflexes. In H. D. Kimmel, E. H. van Olst, & J. F.
Orlebeke (Eds.), The orienting reflex in humans. An international
conference sponsored by the Scientific Affairs Division of the North Atlantic
Treaty Organization (pp. 137-167). Hillsdale, NJ: Lawrence Erlbaum Associates.
Graham, F. K. (1980). Control of reflex blink
excitability. In R. F. Thompson, L. H. Hicks, & V. B. Shvyrkov (Eds.), Neural
mechanisms of goal-directed behavior and learning (pp. 511-519). New York:
Academic Press.
Gratton, G., Coles, M. G. H., & Donchin, E.
(1983). A new method for off-line removal of ocular artifact. Electroencephalography
and Clinical Neurophysiology, 55, 468-484.
Greenhouse, S. W., & Geisser, S. (1959). On
methods in the analysis of profile data. Psychometrika, 24,
95-112.
Groenewegen, H. J., & Berendse, H. W. (1994). The
specificity of the 'nonspecific' midline and intralaminar thalamic nuclei. Trends
in Neuroscience, 17, 52-57.
Hackley, S. A., & Graham, F. K. (1987). Effects of
attending selectively to the spatial position of reflex-eliciting and
reflex-modulating stimuli. Journal of Experimental Psychology: Human
Perception and Performance, 13, 411-424.
Hackley, S. A., Woldorff, M., & Hillyard, S. A.
(1990). Cross-modal selective attention effects on retinal, myogenic, brainstem,
and cerebral evoked potentials. Psychophysiology, 27, 195-208.
Hari, R., Aittoniemi, K., Järvinen, M. L., Katila,
T., & Varpula, T. (1980). Auditory evoked transient and sustained magnetic
fields of the human brain: Localization of neural generators. Experimental
Brain Research, 40, 237-240.
Hari, R., Kaila, K., Katila, T., Tuomisto, T., &
Varpula, T. (1982). Interstimulus interval dependence of the auditory vertex
response and its magnetic counterpart: Implications for their neural generation.
Electroencephalography and Clinical Neurophysiology, 54, 561-569.
Harrison, J. B., Woolf, N. J., & Buchwald, J. S.
(1990). Cholinergic neurons of the feline pontomesencephalon. I. Essential role
in 'wave A' generation. Brain Research, 520, 43-54.
Hashimoto, I., Mashiko, T., Yoshikawa, K., Mizuta, T.,
Imada, T., & Hayashi, M. (1995). Neuromagnetic measurements of the human
primary auditory response. Electroencephalography and Clinical
Neurophysiology, 96, 348-356.
Huttunen, J., Hari, R., Vanni, S. (1987). Crossmodal
interaction is reflected in vertex potentials but not in evoked magnetic fields.
Acta Neurologica Scandinavica, 75, 410-416.
Jasper, H. H. (1958). The ten-twenty electrode system
of the International Federation. Electroencephalography and Clinical
Neurophysiology, 10, 371-375.
Koch, M., & Schnitzler, H-U. (1997). The acoustic
startle response in rats—circuits mediating evocation, inhibition and
potentiation. Behavioural Brain Research, 89, 35-49.
Kodsi, M. H., & Swerdlow, N. R. (1997a).
Regulation of prepulse inhibition by ventral pallidal projections. Brain
Research Bulletin, 43, 219-228.
Kodsi, M. H., & Swerdlow, N. R. (1997b). Reduced
prepulse inhibition after electrolytic lesions of nucleus accumbens subregions
in the rat. Brain Research, 773, 45-52.
Leitner, D. S., Powers, A. S., & Hoffman, H. S.
(1979). The neural system for the inhibition of startle. Bulletin of the
Psychonomic Society, 14, 410-412.
Leitner, D. S., Powers, A. S., Stitt, C. L., &
Hoffman, H. S. (1981). Midbrain reticular formation involvement in the
inhibition of acoustic startle. Physiology and Behavior, 26,
259-268.
Liégeois-Chauvel, C., Musolino, A., & Chauvel, P.
(1991). Localization of the primary auditory area in man. Brain, 114,
139-153.
Liégeois-Chauvel, C., Musolino, A., Badier, J. M.,
Marquis, P., & Chauvel, P. (1994). Evoked potentials recorded from the
auditory cortex in man: Evaluation and topography of the middle latency
components. Electroencephalography and Clinical Neurophysiology, 92,
204-214.
Loveless, N. E., & Brunia, C. H. M. (1990).
Effects of rise-time on late components of the auditory evoked potential. Journal
of Psychophysiology, 4, 369-380.
Loveless, N. E., & Hari, R. (1993). Auditory
evoked fields covary with perceptual grouping. Biological Psychology, 35,
1-15.
Loveless, N., Hari, R., Hämäläinen, M., &
Tiihonen, J. (1989). Evoked responses of human auditory cortex may be enhanced
by preceding stimuli. Electroencephalography and Clinical Neurophysiology,
74, 217-227.
Marks, L. E. (1988). Magnitude estimation and sensory
matching. Perception & Psychophysics, 43, 511-525.
Marks, L. E., Szczesiul, R., & Ohlott, P. (1986).
On the cross-modal perception of intensity. Journal of Experimental
Psychology: Human Perception and Performance, 12, 517-534.
Marks, L. E., & Warner, E. (1991). Slippery
context effect and critical bands. Journal of Experimental Psychology: Human
Perception and Performance, 17, 986-996.
Miller, G. A., Gratton, G., & Yee, C. M. (1989).
Generalized implementation of an eye movement correction procedure. Psychophysiology,
25, 241-243.
Näätänen, R., & Picton, T. (1987). The N1 wave
of the human electric and magnetic response to sound: A review and an analysis
of the component structure. Psychophysiology, 24, 375-425.
Naber, G., Kathman, N., & Engel, R. R. (1992). P50
suppression in normal subjects: Influence of stimulus intensity, test repetition, and
presentation mode. Journal of Psychophysiology, 6, 47-53.
Nagamoto, H. T., Adler, L. E., Waldo, M. C., &
Freedman, R. (1989). Sensory gating in schizophrenics and normal controls:
Effects of changing stimulation interval. Biological Psychiatry, 25,
549-561.
Nagamoto,
H. T., Adler, L. E., Waldo, M. C., Griffith, J., & Freedman, R. (1991).
Gating of auditory response in schizophrenics and normal controls: Effects of
recording site and stimulation interval on the P50 wave. Schizophrenia
Research, 4, 31-40.
Nelson, D. A., & Lassman, F. M. (1968). Effects of
intersignal interval on the human auditory evoked response. Journal of the
Acoustical Society of America, 44, 1529-1532.
Neylan,
T. C., Fletcher, D. J., Lenoci, M., McCallin, K., Weiss, D. S., Schoenfeld, F.B.,
Marmar, C.R., & Fein, G. (1999). Sensory gating in chronic posttraumatic
stress disorder: reduced auditory P50 suppression in combat veterans. Biological
Psychiatry, 46,1656-1664.
Norris,
C. M., & Blumenthal, T. D. (1996). A relationship between inhibition of the
acoustic startle response and the protection of prepulse processing. Psychobiology,
24, 160-168.
Onishi, S., & Davis, H. (1968). Effects of
duration and rise time of tone bursts on evoked V potentials. Journal of the
Acoustical Society of America, 44, 582-591.
Parker, S., & Schneider, B. (1994). The stimulus
range effect: Evidence for top-down control of sensory intensity in audition.
Perception & Psychophysics, 56, 1-11.
Perlstein, W. M., Fiorito, E., Simons, R. F., &
Graham, F. K. (1993). Lead stimulation effects on reflex blink, exogenous brain
potentials, and loudness judgments. Psychophysiology, 30, 347-358.
Perrault, N., & Picton, T. W. (1984).
Event-related potentials recorded from the scalp and nasopharynx. I. N1 and P2. Electroencephalography
and Clinical Neurophysiology, 59, 177-194.
Picton, T. W., Goodman, W. S., & Bryce, D. P.
(1970). Amplitude of evoked responses to tones of high intensity. Acta
Otolaryngology, 70, 77-82.
Picton, T. W., Woods, D. L., & Proulx, G. B.
(1978a). Human auditory sustained potentials. I. The nature of the response. Electroencephalography
and Clinical Neurophysiology, 45, 186-197.
Picton, T. W., Woods, D. L., & Proulx, G. B.
(1978b). Human auditory sustained potentials. II. Stimulus relationships. Electroencephalography
and Clinical Neurophysiology, 45, 198-210.
Putnam, L. E., & Roth, W. T. (1990). Effects of
stimulus repetition, duration, and rise time on startle blink and automatically
elicited P300. Psychophysiology, 27, 275-297.
Rankin, K. M., & Marks, L. E. (1991). Differential
context effects in taste perception. Chemical Senses, 16, 617-629.
Reite, M., Zimmerman, J. T., Edrich, J., &
Zimmerman, J. E. (1982). Auditory evoked magnetic fields: Response amplitude vs.
stimulus intensity. Electroencephalography and Clinical Neurophysiology, 54,
147-152.
Rimpel, J., Geyer, D., & Hopf, H. C. (1982).
Changes in the blink responses to combined trigeminal, acoustic, and visual
repetitive stimulation, studied in the human subject. Electroencephalography
and Clinical Neurophysiology, 54, 552-560.
Roemer, R. A., Shagass, C., & Teyler, T. J.
(1984). Do human evoked potentials habituate? In H. V. S. Peeke & L.
Petrinovich (Eds.), Habituation, sensitization, and behavior (pp.
325-346). New York: Academic Press.
Roth, W. T., Dorato, K. H., & Kopell, B. S.
(1984). Intensity and task effects on evoked physiological responses to noise
bursts. Psychophysiology, 21, 466-481.
Roth, W. T., Krainz, P. L., Ford, J. M., Tinklenberg,
J. R., Rothbart, R. M., & Kopell, B. S. (1976). Parameters of temporal
recovery of the human auditory evoked potential. Electroencephalography and
Clinical Neurophysiology, 40, 623-632.
Rothman, H. H., Davis, H., & Hay, I. S. (1970).
Slow evoked cortical potentials and temporal features of stimulation. Electroencephalography
and Clinical Neurophysiology, 29, 225-232.
Saitoh, K., Tilson, H. A., Shaw, S., & Dyer, R. S.
(1987). Possible role of the brainstem in the mediation of prepulse inhibition
in the rat. Neuroscience Letters, 7, 216-222.
Scherg, M., Hari, R., & Hämäläinen, M. (1989).
Frequency-specific sources of the auditory N19-P30 detected by a multiple source
analysis of evoked magnetic fields and potentials. In S. J. Williamson, M. Hoke,
G. Stroink, & M. Kotani (Eds.), Advances in Biomagnetism (pp.
97-100). New York: Plenum.
Scherg, M., & von Cramon, D. (1986). Evoked dipole
source potentials of the human auditory cortex. Electroencephalography and
Clinical Neurophysiology, 65, 344-360.
Schicatano, E. J., Peshori, K. R., Gopalaswamy, R.,
Sahay, E., & Evinger, C. (2000). Reflex excitability regulates prepulse
inhibition. Journal of Neuroscience, 20, 4240-4247
Schneider, B., & Parker, S. (1990). Does stimulus
context affect loudness or only loudness judgments? Perception &
Psychophysics, 48, 409-418.
Silverstein, L. D., & Graham, F. K. (1978).
Eyeblink EMG: A miniature eyelid electrode for recording from orbicularis oculi.
Psychophysiology, 15, 377-379.
Stevens, S. S. (1956). The direct estimation of
sensory magnitudes-loudness. American Journal of Psychology, 69,
1-25.
Swerdlow, N. R., Geyer, M. A., Blumenthal, T. D.,
& Hartman, P. L. (1999). Effects of discrete acoustic prestimuli on
perceived intensity and behavioral responses to startling acoustic and tactile
stimuli. Psychobiology, 27, 547-556.
Szczepaniak, W. S., & Møller, A. R. (1993).
Interaction between auditory and somatosensory systems:
A study of evoked potentials in the inferior colliculus. Electroencephalography
and Clinical Neurophysiology, 88, 508-515.
Wasman, M., & Gluck, H. (1975). Recovery functions
of somatosensory evoked responses in slow learners. Psychophysiology, 12,
371-376.
Webster, W. R., & Aitkin, L. M. (1971). Evoked
potential and single unit studies of neural mechanisms underlying the effects of
repetitive stimulation in the auditory pathway. Electroencephalography and
Clinical Neurophysiology, 31, 581-592.
Weinberger, N. M., & Diamond, D. M. (1987).
Physiological plasticity in auditory cortex: Rapid induction by learning. Progress
in Neurobiology, 29, 1-55.
Weinberger, N. M., & Diamond, D. M. (1988).
Dynamic modulation of the auditory system by associative learning. In G. M.
Edelman, W. E. Gall, & W. M. Cowan (Eds.), Auditory Function:
Neurobiological Bases of Hearing (pp. 485-512). New York: Wiley.
Woldorff, M. G. (1993). Disortion of ERP averages due
to overlap from temporally adjacent ERPs: Analysis and correction. Psychophysiology,
30, 98-119.
![]()
1. An eight-subject pilot study determined that no acoustic
artifact was detectable. Delivery
of airpuffs that were not directed toward the skin did not produce PPI of blink,
in contrast to the presence of PPI effects when puff was directed toward the
skin. These subjects also reported that they did not detect any puff-related any
acoustic artifact at puff presentation.
2.
EMG was corrected for potential EOG contamination since these data were
recorded from the same mastoid reference used for measuring ERPs, and therefore,
they are subject to the same potential contaminating effects of blinks and eye
movements.
3. Peaks could not be identified in 13% of the total sample,
distributed equally across recording sites 2and conditions 2and unequally across components [c2(3)
= 42.36, p<.005], with points chosen at the window extremes for N19
most frequently and for P50 least frequently.
4. An initial PCA on a data set including the midline N100 and
P200 components yielded compound or biphasic waveforms with each component
having high negative loadings on the other component.
Compound waveforms for these components might be expected given that,
both in the grand-mean ERP waveforms and in the peak scores, N100 and P200 were
affected similarly by the pairing manipulation. As Donchin and Heffley (1978) note, when individual ERP
components are highly correlated across experimental variables and electrodes,
they will produce a loading vector and component scores based on the compound
waveform. Consequently, PCA was not
used to measure the later-latency components midline N100 and P200 and lateral
C1 and C2. Perlstein et al. (1993)
did not conduct a PCA on points beyond 85 ms, but a re-analysis of their data
showed that N100 and P200 also had reciprocal negative loadings in that study.
5. Correction
of these muscle responses for vertical and horizontal EOG had negligible effects
on morphology and amplitude of the initial negative-positive deflections of the
masseter and post-auricularis muscles; effects on frontalis and later portions
of the masseter and post-auricularis were more substantial.
6.
In addition to the primary EOG-corrected ERP data , we also examined ERPs
for additional corrections involving the facial/scalp muscles. These
corrections were conducted by subjecting the EOG-corrected single-trial data to
subsequent steps in the correction algorithm that used the muscle data for
correction. Thus, correction
yielded four additional sets of waveforms: 1) masseter-corrected for EOG then
masseter; 2) post-auricularis-corrected for EOG then post-auricularis; 3)
frontalis-corrected for EOG then medial frontalis; and 4) All-corrected for EOG,
masseter, PAR, and frontalis, in order.
![]()
Figure
1. Waveforms for blink and
scalp/facial EMG responses to single control tones and tones preceded by puffs
at 60- and 360-ms SOA. Facial EMGs
have been corrected for vertical and horizontal eye movements.
(Note ordinate scale differences).
Figure
2. Grand mean ERP waveforms at
midline (Fz, Cz) and lateral (F8, T4/6) recording sites as a function of single
and paired stimulus conditions. (Note ordinate scale differences).
Figure
3. Expanded midline ERP grand mean
waveforms showing midlantency ERP components in the first 85 ms of the recording
epoch.
Figure
4. Factor loadings from the PCA on
the first 85 ms of the midline midlatency ERPs.
Figure
5. Factor loadings from the
“7-lead” PCA on the first 85 ms of midlatency ERP and EMG waveforms.
This PCA included ERPs at all four recording sites (Fz, Cz, F8, T4/6) and
three EMG recording sites (masseter, post-auricularis, medial frontalis).
Figure
6. Mean factor scores from the
“7-lead” PCA on midlatency deflections as a function of pairing condition
and recording site.
Figure
7. Mean range-corrected
base-to-peak amplitudes for midline P30 and P50 (top) and for the three N100
components (bottom) as a function of pairing condition.
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}